‘longevity’ directory
- See Also
- Gwern
- Links
- “Digit Regeneration in Mice Is Stimulated by Sequential Treatment With FGF2 and BMP2”
- “Inside the Stealthy Startup R3 Bio That Pitched Brainless Human Clones: The Ultimate Plan to Live Forever Is a Brand New Body”, Regalado 2026
- “A Billionaire-Backed Startup Wants to Grow ‘Organ Sacks’ to Replace Animal Testing: R3 Bio Has a Bold Idea for Replacing Lab Animals: Genetically-Engineered Whole Organ Systems That Lack a Brain. The Long-Term Goal, Says a Cofounder, Is to Make Human Versions”, Mullin 2026
- “My Hobby: Running Deranged Surveys; You Can Just Ask People Things”, Gao 2026
- “Meet the Vitalists: the Hardcore Longevity Enthusiasts Who Believe Death Is ‘Wrong’; They Argue We Need a Revolution—And More and More Influential Scientists, Funders, and Politicians Are Taking Them Seriously”, Hamzelou 2026
- “Sterilization and Contraception Increase Lifespan across Vertebrates”, Garratt et al 2025
- “Lysergic Acid Diethylamide Extends Lifespan in Caenorhabditis Elegans”, Carrilho et al 2025
- “Cardiorespiratory Fitness in Adolescence and Premature Mortality: Widespread Bias Identified Using Negative Control Outcomes and Sibling Comparisons”, Ballin et al 2025
- “Subtitling Your Life: Hearing Aids and Cochlear Implants Have Been Getting Better for Years, but a New Type of Device—Eyeglasses That Display Real-Time Speech Transcription on Their Lenses—Are a Game-Changing Breakthrough”, Owen 2025
- “Ethically Sourced ‘Spare’ Human Bodies Could Revolutionize Medicine: Human ‘Bodyoids’ Could Reduce Animal Testing, Improve Drug Development, and Alleviate Organ Shortages”, Charlesworth et al 2025
- “How Bryan Johnson, Who Wants to Live Forever, Sought Control via Confidentiality Agreements: That Control Is Starting to Fray As Johnson, a Longevity Guru Known for Performing Experiments on His Body, Faces Backlash”, Grind 2025
- “Replacement As an Aging Intervention”, Lore et al 2025
- “No Effect of Additional Education on Long-Term Brain Structure—A Preregistered Natural Experiment in Thousands of Individuals”, Judd & Kievit 2025
- “No Effect of Education on Telomere Length: a Natural Experiment in Aging Individuals”, Judd & Kievit 2025
- “Crypto Millionaire Fuels Push to Transform Brain Research: James Fickel Has Dedicated $200 Million He Made Betting on Ether to Becoming One of the World’s Biggest Investors in Those Fields”, Vance 2024
- “(Inaccurate) Beliefs about Skill Decay”, Connolly et al 2024
- “Pervasive Findings of Directional Selection Realize the Promise of Ancient DNA to Elucidate Human Adaptation”, Akbari et al 2024
- “The Global Pattern of Centenarians Highlights Deep Problems in Demography”, Newman 2024
- “Spatial Cognitive Ability Is Associated With Longevity in Food-Caching Chickadees”, Welklin et al 2024
- “The Lifetime Costs of Bad Health”, Nardi et al 2024
- “Looks and Longevity: Do Prettier People Live Longer?”, Sheehan & Hamermesh 2024
- “Unveiling the Epigenetic Impact of Vegan versus Omnivorous Diets on Aging: Insights from the Twins Nutrition Study (TwiNS)”, Dwaraka et al 2023
- “Who Is Bryan Johnson’s Doctor? Meet Oliver Zolman”, Mikhail 2023
- “Somatic Mutations in Human Aging: New Insights from DNA Sequencing and Inherited Mutations”, Chatsirisupachai & Magalhães 2023
- “Longevity Factor Klotho Enhances Cognition in Aged Nonhuman Primates”, Castner et al 2023
- “Premature Aging and Reduced Cancer Incidence Associated With Near-Complete Body-Wide Myc Inactivation”, Wang et al 2023
- “Late-Life Rapamycin Treatment Enhances Cardiomyocyte Relaxation Kinetics and Reduces Myocardial Stiffness”, Chakraborty et al 2023
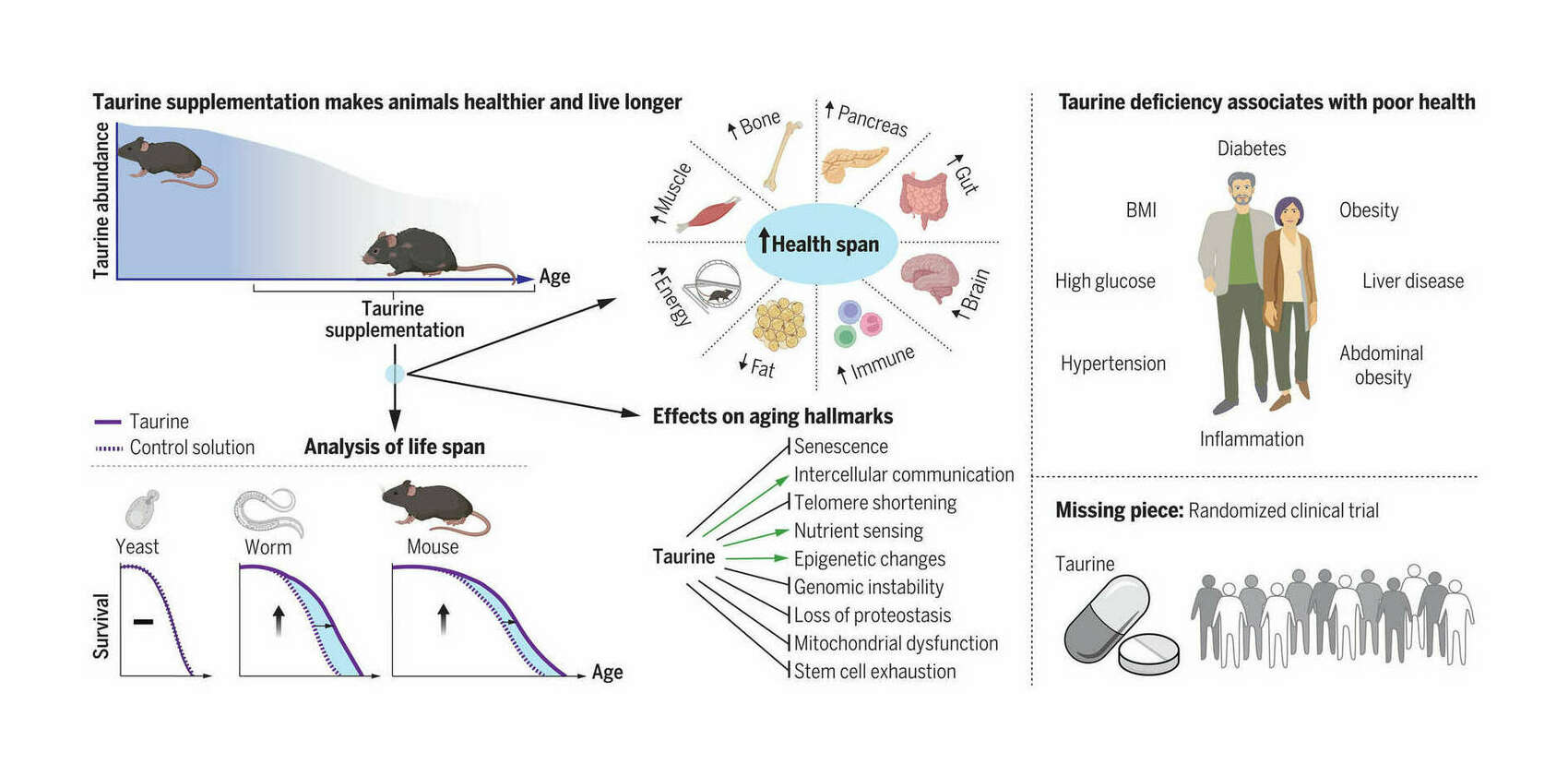
- “Taurine Deficiency As a Driver of Aging”, Singh et al 2023
- “Using Sequences of Life-Events to Predict Human Lives”, Savcisens et al 2023
- “Mendelian Randomization Supports Causality between Overweight Status and Accelerated Aging”, Chen et al 2023
- “DNA Repair and Anti-Cancer Mechanisms in the Longest-Living Mammal: the Bowhead Whale”, Firsanov et al 2023
- “Intelligence and Life Expectancy in Late Adulthood: A Meta-Analysis”, Sanchez-Izquierdo et al 2023
- “Age-Dependent Topic Modeling of Comorbidities in UK Biobank Identifies Disease Subtypes With Differential Genetic Risk”, Jiang et al 2023
- “Evidence for the Role of Selection for Reproductively Advantageous Alleles in Human Aging”, Long & Zhang 2023
- “Hypoxia Extends Lifespan and Neurological Function in a Mouse Model of Aging”, Rogers et al 2023
- “Five Years Later, With Double the Demographic Data, Naked Mole-Rat Mortality Rates Continue to Defy Gompertzian Laws by Not Increasing With Age”, Ruby et al 2023
- “Sam Altman Invested $180 Million into a Company Trying to Delay Death: Can Anti-Aging Breakthroughs Add 10 Healthy Years to the Human Life Span? The CEO of OpenAI Is Paying to Find Out”, Regalado 2023
- “Loss of Smelling Is an Early Marker of Aging and Is Associated With Inflammation and DNA Damage in C57BL/6J Mice”, Dan et al 2023
- “Mortality Postponement and Compression at Older Ages in Human Cohorts”, McCarthy & Wang 2023
- “Status and Mortality: Is There a Whitehall Effect in the United States?”, Nicholas 2023
- “Effect of Basal Metabolic Rate on Lifespan: a Sex-Specific Mendelian Randomization Study”, Ng & Schooling 2023
- “Cell Tree Rings: the Shape of Somatic Evolution As a Human Aging Timer”, Csordas et al 2022
- “Rejuvenating Senescent Cells and Organisms With Only Ultrasound”, Kumar et al 2022
- “Today’s Older Adults Are Cognitively Fitter Than Older Adults Were 20 Years Ago, but When and How They Decline Is No Different Than in the Past”, Gerstorf et al 2022
- “Lifespan Benefits for the Combination of Rapamycin plus Acarbose and for Captopril in Genetically Heterogeneous Mice”, Strong et al 2022
- “A Cocktail of Rapamycin, Acarbose and Phenylbutyrate Prevents Age-Related Cognitive Decline in Mice by Altering Aging Pathways”, Jiang et al 2022
- “Old Plasma Dilution Reduces Human Biological Age: a Clinical Study”, Kim et al 2022
- “Slow and Negligible Senescence among Testudines Challenges Evolutionary Theories of Senescence”, Silva et al 2022
- “A Hierarchical Process Model Links Behavioral Aging and Lifespan in C. Elegans”, Oswal et al 2022
- “Exercise Molecule Burns Away Hunger”, Kim & Sternson 2022
- “An Exercise-Inducible Metabolite That Suppresses Feeding and Obesity”, Li et al 2022
- “An Explanation for Negligible Senescence in Animals”, Xia & Møller 2022
- “Clonal Dynamics of Hematopoiesis across the Human Lifespan”, Mitchell et al 2022
- “Mendelian Randomization of Genetically Independent Aging Phenotypes Identifies LPA and VCAM1 As Biological Targets for Human Aging”, Timmers et al 2022
- “Rare Genetic Variants Correlate With Better Processing Speed”, Song et al 2022
- “Longevity, Cellular Senescence and the Gut Microbiome: Lessons to Be Learned from Crocodiles”, Siddiqui et al 2021
- “Coevolution of Brain Size and Longevity in Parrots”, Smeele et al 2021
- “Polygenic Basis and Biomedical Consequences of Telomere Length Variation”, Codd et al 2021
- “Escape of Hair Follicle Stem Cells Causes Stem Cell Exhaustion during Aging”, Zhang et al 2021
- “Increased Somatic Mutation Burdens in Normal Human Cells due to Defective DNA Polymerases”, Robinson et al 2021
- “Human Mortality at Extreme Age”, Belzile et al 2021
- “Lithium Can Mildly Increase Health during Aging but Not Lifespan in Mice”, Nespital et al 2021
- “Extending Human Healthspan and Longevity: a Symposium Report”, DeVito et al 2021
- “Somatic Mutation Rates Scale With Lifespan across Mammals”, Cagan et al 2021
- “Counteracting Age-Related VEGF Signaling Insufficiency Promotes Healthy Aging and Extends Life Span”, Grunewald et al 2021
- “An Aged Immune System Drives Senescence and Aging of Solid Organs”, Yousefzadeh et al 2021
- “Beneficial and Detrimental Effects of Reactive Oxygen Species on Lifespan: A Comprehensive Review of Comparative and Experimental Studies”, Shields et al 2021
- “The Distribution of Cellular Turnover in the Human Body”, Sender & Milo 2021
- “Carbohydrate Restriction for Diabetes: Rediscovering Centuries-Old Wisdom”, Lennerz et al 2021
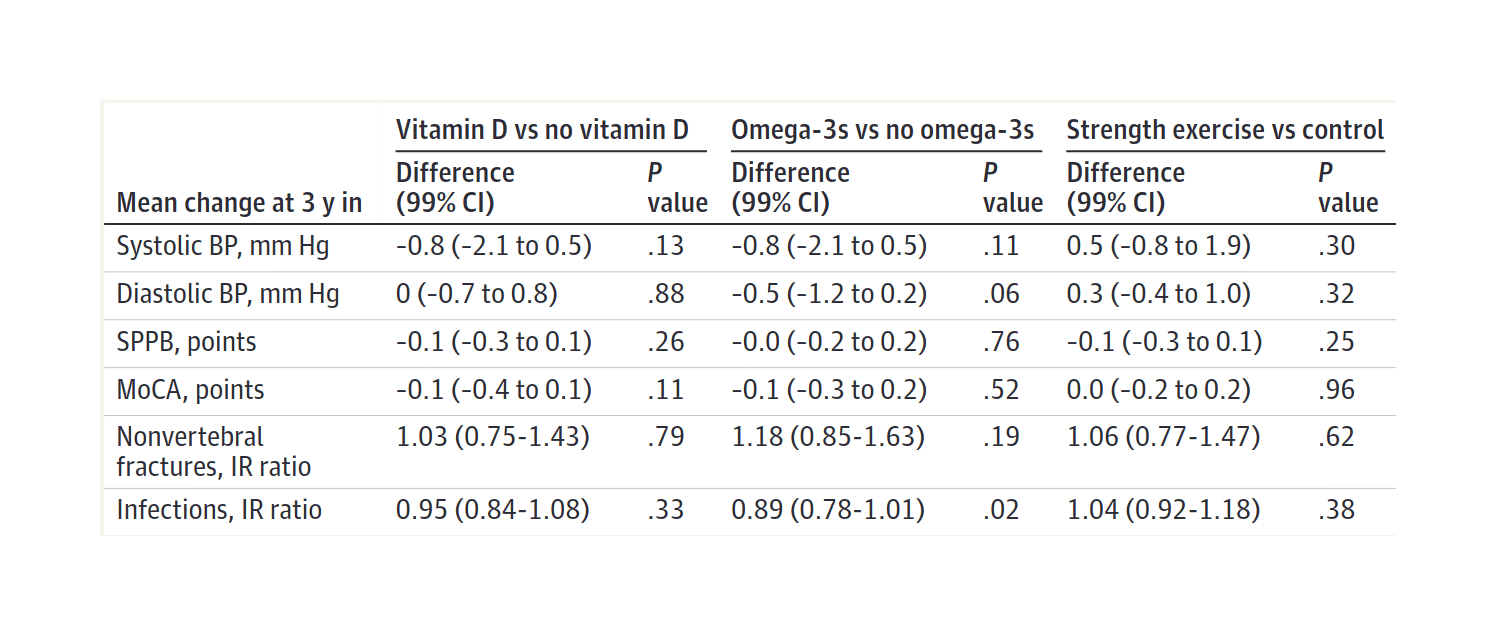
- “Effect of Vitamin D Supplementation, Omega-3 Fatty Acid Supplementation, or a Strength-Training Exercise Program on Clinical Outcomes in Older Adults: The DO-HEALTH Randomized Clinical Trial”, Bischoff-Ferrari et al 2020
- “Surprisingly Long Survival of Premature Conclusions about Naked Mole-Rat Biology”, Braude et al 2020
- “C60 in Olive Oil Causes Light-Dependent Toxicity and Does Not Extend Lifespan in Mice”, Grohn et al 2020
- “CRISPR-Enhanced Human Adipocyte ‘Browning’ As Cell Therapy for Metabolic Disease”, Tsagkaraki et al 2020
- “The Burden of Rare Protein-Truncating Genetic Variants on Human Lifespan”, Liu et al 2020
- “Supercentenarian and Remarkable Age Records Exhibit Patterns Indicative of Clerical Errors and Pension Fraud”, Newman 2020
- “Was Jeanne Calment the Oldest Person Who Ever Lived—Or a Fraud? Some Researchers Have Cast Doubt on the Record of the Celebrated Supercentenarian”, Collins 2020
- “Cryonics for All?”, Thau 2020
- “Vitamin D Supplementation for Prevention of Mortality in Adults”, Bjelakovic et al 2019b
- “A Single Combination Gene Therapy Treats Multiple Age-Related Diseases”, Davidsohn et al 2019
- “The Genetics of Human Aging”, Melzer et al 2019
- “Effect of Lower Versus Higher Red Meat Intake on Cardiometabolic and Cancer Outcomes: A Systematic Review of Randomized Trials”, Zeraatkar et al 2019
- “Exercise Conditioned Plasma Dampens Inflammation via Clusterin and Boosts Memory”, Miguel et al 2019
- “A Prospective Analysis of Genetic Variants Associated With Human Lifespan”, Wright et al 2019
- “Restoration of Brain Circulation and Cellular Functions Hours Post-Mortem”, Vrselja et al 2019
- “Genetically Heterogeneous Mice Exhibit a Female Survival Advantage That Is Age-Specific and Site-Specific: Results from a Large Multi-Site Study”, Cheng et al 2019
- “Relaxed Selection Limits Lifespan by Increasing Mutation Load”, Cui et al 2019
- “Turning Back Time With Emerging Rejuvenation Strategies”, Mahmoudi et al 2019
- “An Evolutionary Perspective on Why Food Overconsumption Impairs Cognition”, Mattson 2019
- “A Longitudinal Big Data Approach for Precision Health”, Rose et al 2019
- “What Do We Need to Know to Treat Degenerative Aging As a Medical Condition to Extend Healthy Lifespan?”, Stambler 2019
- “Effects of Intermittent Fasting on Health, Aging, and Disease”, Cabo & Mattson 2019
- “Evidence That Jeanne Calment Died in 1934-Not 1997”, Zak 2019
- “Genomic Underpinnings of Lifespan Allow Prediction and Reveal Basis in Modern Risks”, Timmers et al 2018
- “The Protective Effect of Melatonin Against Age-Associated, Sarcopenia-Dependent Tubular Aggregate Formation, Lactate Depletion, and Mitochondrial Changes”, Sayed et al 2018
- “Anti-Aging Food That Improves Markers of Health in Senior Dogs by Modulating Gut Microbiota and Metabolite Profiles”, Gebreselassie et al 2018
- “Quantitative Analysis of Population-Scale Family Trees With Millions of Relatives”, Kaplanis et al 2018
- “What Science Is like in North Korea: Isolated from the Rest of the World, North Korean Researchers Struggle to Balance Rigorous Scientific Work With the Demands of a Dictator”, Fiscutean 2018
- “Effect of Low-Fat vs Low-Carbohydrate Diet on 12-Month Weight Loss in Overweight Adults and the Association With Genotype Pattern or Insulin SecretionThe DIETFITS Randomized Clinical Trial”, Association 2018
- “Autophagy and the Cell Biology of Age-Related Disease”, Leidal et al 2018
- “Early Time-Restricted Feeding Improves Insulin Sensitivity, Blood Pressure, and Oxidative Stress Even without Weight Loss in Men With Prediabetes”, Sutton et al 2018
- “Selection for Long and Short Sleep Duration in Drosophila Melanogaster Reveals the Complex Genetic Network Underlying Natural Variation in Sleep”, Harbison et al 2017
- “Genome-Wide Meta-Analysis Associates HLA-DQA1/DRB1 and LPA and Lifestyle Factors With Human Longevity”, Joshi et al 2017
- “A Long Journey to Reproducible Results: Replicating Our Work Took Four Years and 100,000 Worms but Brought Surprising Discoveries”, Lithgow et al 2017
- “Identifying Genetic Variants That Affect Viability in Large Cohorts”, Mostafavi et al 2017
- “Genetic Contribution to Two Factors of Neuroticism Is Associated With Affluence, Better Health, and Longer Life”, Hill et al 2017
- “Safe Landing Strategies During a Fall: Systematic Review and Meta-Analysis”, Moon & Sosnoff 2017
- “Dietary Restriction and AMPK Increase Lifespan via Mitochondrial Network and Peroxisome Remodeling”, Weir et al 2017
- “2,4-Dinitrophenol, the Inferno Drug: a Netnographic Study of User Experiences in the Quest for Leanness”, McVeigh et al 2017
- “Nutritional Ecology and the Evolution of Aging”, Raubenheimer et al 2016
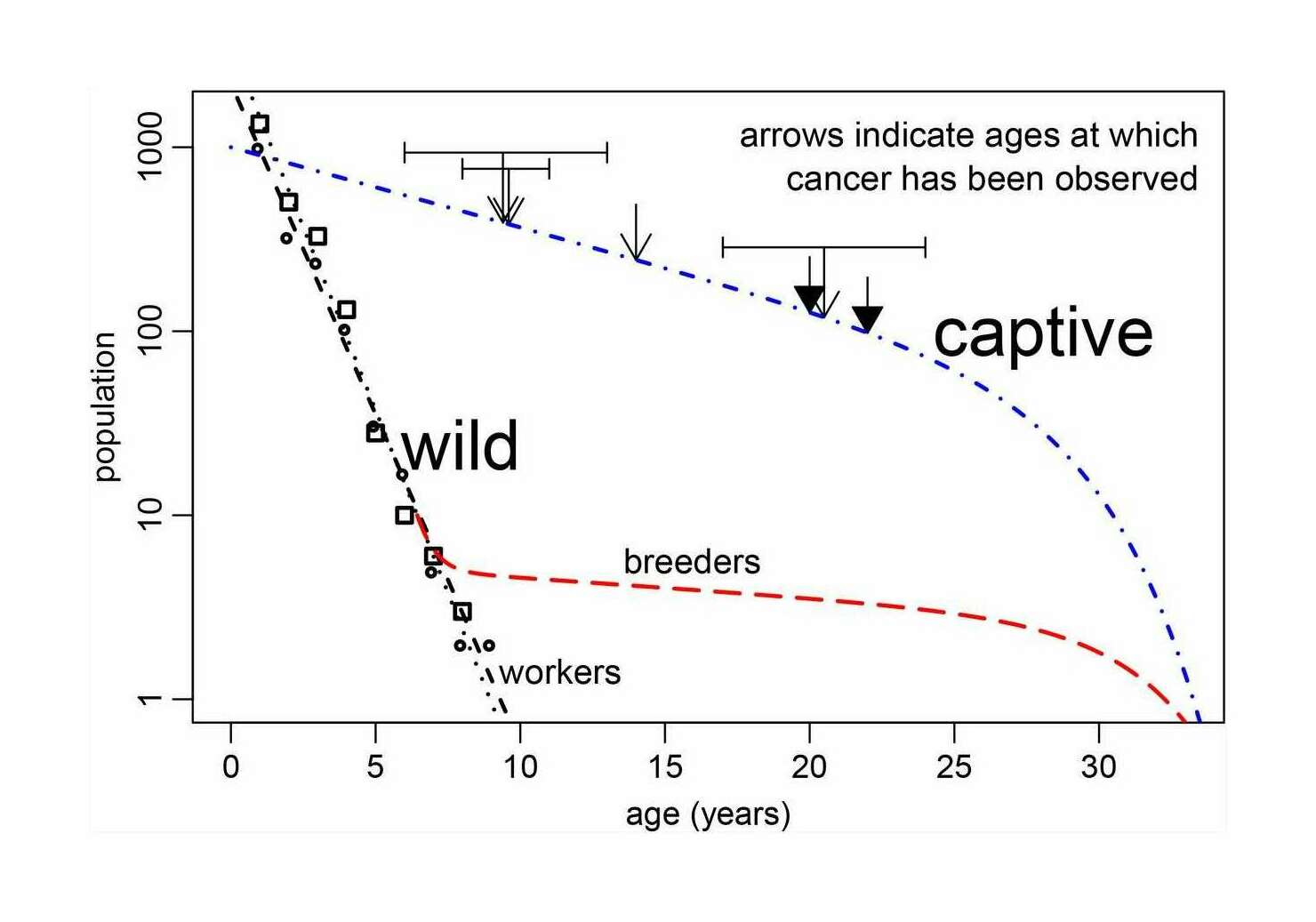
- “A Hypothesis to Explain Cancers in Confined Colonies of Naked Mole Rats”, Hochberg et al 2016
- “Transient Rapamycin Treatment Can Increase Lifespan and Healthspan in Middle-Aged Mice”, Bitto et al 2016
- “Performance of Informative Priors Skeptical of Large Treatment Effects in Clinical Trials: A Simulation Study”, Pedroza et al 2015
- “The Tail End”, waitbutwhy 2015
- “Consciousness without Cortex: a Hydranencephaly Family Survey”, Aleman & Merker 2014
- “Vitamin D and Vitamin D Analogues for Preventing Fractures in Post-Menopausal Women and Older Men”, Avenell et al 2014
- “Vitamin D Supplementation for Prevention of Cancer in Adults (Review)”, G et al 2014
- “A Review of Creatine Supplementation in Age-Related Diseases: More Than a Supplement for Athletes”, Smith et al 2014
- “Who By Very Slow Decay”, Alexander 2013
- “Reproductive Capability Is Associated With Lifespan and Cause of Death in Companion Dogs”, Hoffman et al 2013
- “Multivitamin-Multimineral Supplementation and Mortality: a Meta-Analysis of Randomized Controlled Trials”, Macpherson et al 2013
- “The Caenorhabditis Elegans Lifespan Machine”, Stroustrup et al 2013
- “‘Life in a Germ-Free World’: Isolating Life from the Laboratory Animal to the Bubble Boy § Germ-Free Benefits”, Kirk 2012 (page 7)
- “Terminal Dedifferentiation of Cognitive Abilities”, Wilson et al 2012
- “A Brief History of Cross-Species Organ Transplantation”, Cooper 2012
- “Sitting Time and All-Cause Mortality Risk in 222,497 Australian Adults”, Ploeg 2012
- “Melatonin in Aging and Disease -Multiple Consequences of Reduced Secretion, Options and Limits of Treatment”, Hardeland 2012
- “Happy Aged People Are All Alike, While Every Unhappy Aged Person Is Unhappy in Its Own Way”, Tumminello et al 2011
- “Vitamin D and Cardiovascular Outcomes: A Systematic Review and Meta-Analysis”, Elamin et al 2011
- “Oral Anti-Diabetic Drugs for the Prevention of Type 2 Diabetes”, Phung 2011
- “Intelligence in Youth and All-Cause-Mortality: Systematic Review With Meta-Analysis”, Calvin et al 2011
- “Compression of Morbidity 1980–2011: a Focused Review of Paradigms and Progress”, Fries et al 2011
- “A Genome-Wide Association Study of Aging”, Walter et al 2011
- “Association of Circulating Visfatin Concentrations With Insulin Resistance and Low-Grade Inflammation After Dietary Energy Restriction in Spanish Obese Non-Diabetic Women: Role of Body Composition Changes”, Agueda et al 2010
- “Japan, Checking on Its Oldest, Finds Many Gone”
- “Demographic Consequences of Defeating Aging”, Gavrilov & Gavrilova 2010
- “Biodemography of Human Aging”, Vaupel 2010
- “Typologies of Extreme Longevity Myths”, Young et al 2010
- “Ageing and Its Implications”, Jayanthi et al 2010
- “Plasma Vitamin D and Mortality in Older Men: a Community-Based Prospective Cohort Study”, Michaëlsson 2010
- “A Weak Link in Metabolism: the Metabolic Capacity for Glycine Biosynthesis Does Not Satisfy the Need for Collagen Synthesis”, Meléndez-Hevia et al 2009
- “Good Semen Quality and Life Expectancy: A Cohort Study of 43,277 Men”, Jensen et al 2009
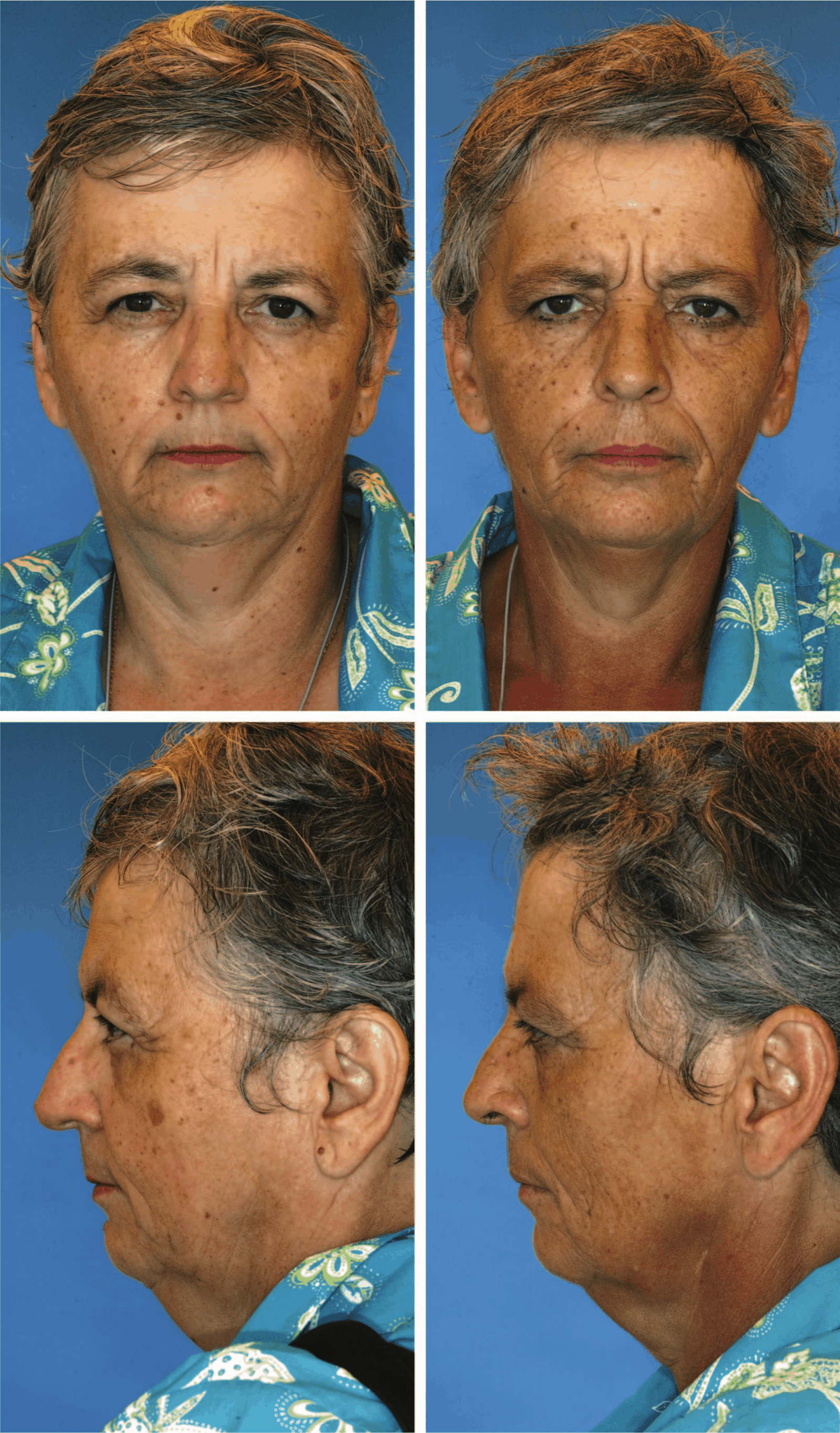
- “Factors Contributing to the Facial Aging of Identical Twins”, Guyuron et al 2009
- “Use of Supplements of Multivitamins, Vitamin C, and Vitamin E in Relation to Mortality”, Pocobelli 2009
- “Vitamin D and Aging”, Tuohimaa 2009 (page 2)
- “Ageing Populations: the Challenges Ahead”, Christensen et al 2009
- “Effects of Calcium and Vitamin D Supplementation on Hip Bone Mineral Density and Calcium-Related Analytes in Elderly Ambulatory Australian Women: A Five-Year Randomized Controlled Trial”, Zhu et al 2008
- “Prolonged Survival to Adulthood of a [22yo] Individual With Hydranencephaly”, Bae et al 2008
- “Influence of Environmental Factors on Facial Aging”, Rexbye et al 2006
- “Understanding the Odd Science of Aging”, Kirkwood 2005
- “A Randomized, Controlled Trial of Quadriceps Resistance Exercise and Vitamin D in Frail Older People: The Frailty Interventions Trial in Elderly Subjects (FITNESS)”, Latham et al 2003
- “The Reserve-Capacity Hypothesis: Evolutionary Origins and Modern Implications of the Trade-Off between Tumor-Suppression and Tissue-Repair”, Weinstein & Ciszek 2002
- “Book Review: Validation of Exceptional Longevity, by Bernard Jeune and James W. Vaupel”, Gavrilov & Gavrilova 2000
- “Prevention of Femoral and Lumbar Bone Loss With Hormone Replacement Therapy and Vitamin D3 in Early Postmenopausal Women: A Population-Based 5–Year Randomized Trial”, Komulainen et al 1999
- “A C. Elegans Mutant That Lives Twice As Long As Wild Type”, Kenyon et al 1993
- Pritikin: The Man Who Healed America's Heart, Monte & Pritikin 1988
- Disguised, Moore & Conn 1985
- “Eternal Youth Kills Ants”, Times 1982
- “What Difference Would It Make If Cancer Were Eradicated? An Examination of the Taeuber Paradox”, Keyfitz 1977
- “Calmly We Walk through This April’s Day”, Schwartz 1967
- “Aging in Germ-Free Mice: Life Tables and Lesions Observed at Natural Death”, Gordon et al 1966
- “Influence Of The Normal Flora On Mucosal Morphology And Cellular Renewal In The Ileum. A Comparison Of Germ-Free And Conventional Mice”, Abrams et al 1963
- “Three Jubilees: L. Tolstoy, Hen. Ibsen, N. Fedorov”
- “The Machine Man of Ardathia”, Flagg 1927
- “Spring and Fall”, Hopkins 1880
- “The Value of Health and Longevity”
- “The Old Fools”, Larkin 2026
- “Association of Neurocognitive and Physical Function With Gait Speed in Midlife Neurology”
- “Nintil”, Ricon 2026
- “Low-Dose Lithium Uptake Promotes Longevity in Humans and Metazoans”
- “Eulogy to the Obits”, Balwit 2026
- “Your Book Review: The Family That Couldn’t Sleep”
- “More Than 230,000 Japanese Centenarians ‘Missing’”
- “Computational Analysis of Lifespan Experiment Reproducibility”
- “Core Pathways of Aging”
- “Shinya Yamanaka – Biographical”
- “James F. Fries, Who Studied the Good Life and How to Live It, Dies at 83”
- “Dietary Magnesium Intake Is Inversely Associated With Mortality in Adults at High Cardiovascular Disease Risk”
- SkiFree, Munroe 2026
- “J’accuse…! Why Jeanne Calment’s 122-Year Old Longevity Record May Be Fake”
- “More Evidence for Jeanne Calment’s Identity Theft Hypothesis”
- Sort By Magic
- Wikipedia (8)
- Miscellaneous
- Bibliography
See Also
Gwern
“Why Correlation Usually ≠ Causation”, Gwern 2014
“The Replication Crisis: Flaws in Mainstream Science”, Gwern 2010
Links
“Digit Regeneration in Mice Is Stimulated by Sequential Treatment With FGF2 and BMP2”
Digit regeneration in mice is stimulated by sequential treatment with FGF2 and BMP2
“Inside the Stealthy Startup R3 Bio That Pitched Brainless Human Clones: The Ultimate Plan to Live Forever Is a Brand New Body”, Regalado 2026
“A Billionaire-Backed Startup Wants to Grow ‘Organ Sacks’ to Replace Animal Testing: R3 Bio Has a Bold Idea for Replacing Lab Animals: Genetically-Engineered Whole Organ Systems That Lack a Brain. The Long-Term Goal, Says a Cofounder, Is to Make Human Versions”, Mullin 2026
“My Hobby: Running Deranged Surveys; You Can Just Ask People Things”, Gao 2026
My hobby: running deranged surveys; you can just ask people things
“Meet the Vitalists: the Hardcore Longevity Enthusiasts Who Believe Death Is ‘Wrong’; They Argue We Need a Revolution—And More and More Influential Scientists, Funders, and Politicians Are Taking Them Seriously”, Hamzelou 2026
“Sterilization and Contraception Increase Lifespan across Vertebrates”, Garratt et al 2025
Sterilization and contraception increase lifespan across vertebrates
“Lysergic Acid Diethylamide Extends Lifespan in Caenorhabditis Elegans”, Carrilho et al 2025
Lysergic Acid Diethylamide extends lifespan in Caenorhabditis elegans
“Cardiorespiratory Fitness in Adolescence and Premature Mortality: Widespread Bias Identified Using Negative Control Outcomes and Sibling Comparisons”, Ballin et al 2025
“Subtitling Your Life: Hearing Aids and Cochlear Implants Have Been Getting Better for Years, but a New Type of Device—Eyeglasses That Display Real-Time Speech Transcription on Their Lenses—Are a Game-Changing Breakthrough”, Owen 2025
“Ethically Sourced ‘Spare’ Human Bodies Could Revolutionize Medicine: Human ‘Bodyoids’ Could Reduce Animal Testing, Improve Drug Development, and Alleviate Organ Shortages”, Charlesworth et al 2025
“How Bryan Johnson, Who Wants to Live Forever, Sought Control via Confidentiality Agreements: That Control Is Starting to Fray As Johnson, a Longevity Guru Known for Performing Experiments on His Body, Faces Backlash”, Grind 2025
“Replacement As an Aging Intervention”, Lore et al 2025
“No Effect of Additional Education on Long-Term Brain Structure—A Preregistered Natural Experiment in Thousands of Individuals”, Judd & Kievit 2025
“No Effect of Education on Telomere Length: a Natural Experiment in Aging Individuals”, Judd & Kievit 2025
No effect of education on Telomere Length: a natural experiment in aging individuals
“Crypto Millionaire Fuels Push to Transform Brain Research: James Fickel Has Dedicated $200 Million He Made Betting on Ether to Becoming One of the World’s Biggest Investors in Those Fields”, Vance 2024
“(Inaccurate) Beliefs about Skill Decay”, Connolly et al 2024
“Pervasive Findings of Directional Selection Realize the Promise of Ancient DNA to Elucidate Human Adaptation”, Akbari et al 2024
“The Global Pattern of Centenarians Highlights Deep Problems in Demography”, Newman 2024
The global pattern of centenarians highlights deep problems in demography
“Spatial Cognitive Ability Is Associated With Longevity in Food-Caching Chickadees”, Welklin et al 2024
Spatial cognitive ability is associated with longevity in food-caching chickadees
“The Lifetime Costs of Bad Health”, Nardi et al 2024
“Looks and Longevity: Do Prettier People Live Longer?”, Sheehan & Hamermesh 2024
“Unveiling the Epigenetic Impact of Vegan versus Omnivorous Diets on Aging: Insights from the Twins Nutrition Study (TwiNS)”, Dwaraka et al 2023
“Who Is Bryan Johnson’s Doctor? Meet Oliver Zolman”, Mikhail 2023
“Somatic Mutations in Human Aging: New Insights from DNA Sequencing and Inherited Mutations”, Chatsirisupachai & Magalhães 2023
Somatic mutations in human aging: New insights from DNA sequencing and inherited mutations
“Longevity Factor Klotho Enhances Cognition in Aged Nonhuman Primates”, Castner et al 2023
Longevity factor klotho enhances cognition in aged nonhuman primates
“Premature Aging and Reduced Cancer Incidence Associated With Near-Complete Body-Wide Myc Inactivation”, Wang et al 2023
“Late-Life Rapamycin Treatment Enhances Cardiomyocyte Relaxation Kinetics and Reduces Myocardial Stiffness”, Chakraborty et al 2023
“Taurine Deficiency As a Driver of Aging”, Singh et al 2023
“Using Sequences of Life-Events to Predict Human Lives”, Savcisens et al 2023
“Mendelian Randomization Supports Causality between Overweight Status and Accelerated Aging”, Chen et al 2023
Mendelian Randomization supports causality between overweight status and accelerated aging
“DNA Repair and Anti-Cancer Mechanisms in the Longest-Living Mammal: the Bowhead Whale”, Firsanov et al 2023
DNA repair and anti-cancer mechanisms in the longest-living mammal: the bowhead whale
“Intelligence and Life Expectancy in Late Adulthood: A Meta-Analysis”, Sanchez-Izquierdo et al 2023
Intelligence and life expectancy in late adulthood: A meta-analysis
“Age-Dependent Topic Modeling of Comorbidities in UK Biobank Identifies Disease Subtypes With Differential Genetic Risk”, Jiang et al 2023
“Evidence for the Role of Selection for Reproductively Advantageous Alleles in Human Aging”, Long & Zhang 2023
Evidence for the role of selection for reproductively advantageous alleles in human aging
“Hypoxia Extends Lifespan and Neurological Function in a Mouse Model of Aging”, Rogers et al 2023
Hypoxia extends lifespan and neurological function in a mouse model of aging
“Five Years Later, With Double the Demographic Data, Naked Mole-Rat Mortality Rates Continue to Defy Gompertzian Laws by Not Increasing With Age”, Ruby et al 2023
“Sam Altman Invested $180 Million into a Company Trying to Delay Death: Can Anti-Aging Breakthroughs Add 10 Healthy Years to the Human Life Span? The CEO of OpenAI Is Paying to Find Out”, Regalado 2023
“Loss of Smelling Is an Early Marker of Aging and Is Associated With Inflammation and DNA Damage in C57BL/6J Mice”, Dan et al 2023
“Mortality Postponement and Compression at Older Ages in Human Cohorts”, McCarthy & Wang 2023
Mortality postponement and compression at older ages in human cohorts
“Status and Mortality: Is There a Whitehall Effect in the United States?”, Nicholas 2023
Status and mortality: Is there a Whitehall effect in the United States?
“Effect of Basal Metabolic Rate on Lifespan: a Sex-Specific Mendelian Randomization Study”, Ng & Schooling 2023
Effect of basal metabolic rate on lifespan: a sex-specific Mendelian Randomization study
“Cell Tree Rings: the Shape of Somatic Evolution As a Human Aging Timer”, Csordas et al 2022
Cell Tree Rings: the shape of somatic evolution as a human aging timer
“Rejuvenating Senescent Cells and Organisms With Only Ultrasound”, Kumar et al 2022
Rejuvenating Senescent Cells and Organisms with Only Ultrasound
“Today’s Older Adults Are Cognitively Fitter Than Older Adults Were 20 Years Ago, but When and How They Decline Is No Different Than in the Past”, Gerstorf et al 2022
“Lifespan Benefits for the Combination of Rapamycin plus Acarbose and for Captopril in Genetically Heterogeneous Mice”, Strong et al 2022
“A Cocktail of Rapamycin, Acarbose and Phenylbutyrate Prevents Age-Related Cognitive Decline in Mice by Altering Aging Pathways”, Jiang et al 2022
“Old Plasma Dilution Reduces Human Biological Age: a Clinical Study”, Kim et al 2022
Old plasma dilution reduces human biological age: a clinical study
“Slow and Negligible Senescence among Testudines Challenges Evolutionary Theories of Senescence”, Silva et al 2022
Slow and negligible senescence among testudines challenges evolutionary theories of senescence
“A Hierarchical Process Model Links Behavioral Aging and Lifespan in C. Elegans”, Oswal et al 2022
A hierarchical process model links behavioral aging and lifespan in C. elegans
“Exercise Molecule Burns Away Hunger”, Kim & Sternson 2022
“An Exercise-Inducible Metabolite That Suppresses Feeding and Obesity”, Li et al 2022
An exercise-inducible metabolite that suppresses feeding and obesity
“An Explanation for Negligible Senescence in Animals”, Xia & Møller 2022
“Clonal Dynamics of Hematopoiesis across the Human Lifespan”, Mitchell et al 2022
“Mendelian Randomization of Genetically Independent Aging Phenotypes Identifies LPA and VCAM1 As Biological Targets for Human Aging”, Timmers et al 2022
“Rare Genetic Variants Correlate With Better Processing Speed”, Song et al 2022
Rare Genetic Variants Correlate with Better Processing Speed
“Longevity, Cellular Senescence and the Gut Microbiome: Lessons to Be Learned from Crocodiles”, Siddiqui et al 2021
Longevity, cellular senescence and the gut microbiome: lessons to be learned from crocodiles
“Coevolution of Brain Size and Longevity in Parrots”, Smeele et al 2021
“Polygenic Basis and Biomedical Consequences of Telomere Length Variation”, Codd et al 2021
Polygenic basis and biomedical consequences of telomere length variation
“Escape of Hair Follicle Stem Cells Causes Stem Cell Exhaustion during Aging”, Zhang et al 2021
Escape of hair follicle stem cells causes stem cell exhaustion during aging
“Increased Somatic Mutation Burdens in Normal Human Cells due to Defective DNA Polymerases”, Robinson et al 2021
Increased somatic mutation burdens in normal human cells due to defective DNA polymerases
“Human Mortality at Extreme Age”, Belzile et al 2021
“Lithium Can Mildly Increase Health during Aging but Not Lifespan in Mice”, Nespital et al 2021
Lithium can mildly increase health during aging but not lifespan in mice
“Extending Human Healthspan and Longevity: a Symposium Report”, DeVito et al 2021
Extending human healthspan and longevity: a symposium report
“Somatic Mutation Rates Scale With Lifespan across Mammals”, Cagan et al 2021
“Counteracting Age-Related VEGF Signaling Insufficiency Promotes Healthy Aging and Extends Life Span”, Grunewald et al 2021
Counteracting age-related VEGF signaling insufficiency promotes healthy aging and extends life span
“An Aged Immune System Drives Senescence and Aging of Solid Organs”, Yousefzadeh et al 2021
An aged immune system drives senescence and aging of solid organs
“Beneficial and Detrimental Effects of Reactive Oxygen Species on Lifespan: A Comprehensive Review of Comparative and Experimental Studies”, Shields et al 2021
“The Distribution of Cellular Turnover in the Human Body”, Sender & Milo 2021
“Carbohydrate Restriction for Diabetes: Rediscovering Centuries-Old Wisdom”, Lennerz et al 2021
Carbohydrate restriction for diabetes: rediscovering centuries-old wisdom
“Effect of Vitamin D Supplementation, Omega-3 Fatty Acid Supplementation, or a Strength-Training Exercise Program on Clinical Outcomes in Older Adults: The DO-HEALTH Randomized Clinical Trial”, Bischoff-Ferrari et al 2020
“Surprisingly Long Survival of Premature Conclusions about Naked Mole-Rat Biology”, Braude et al 2020
Surprisingly long survival of premature conclusions about naked mole-rat biology
“C60 in Olive Oil Causes Light-Dependent Toxicity and Does Not Extend Lifespan in Mice”, Grohn et al 2020
C60 in olive oil causes light-dependent toxicity and does not extend lifespan in mice
“CRISPR-Enhanced Human Adipocyte ‘Browning’ As Cell Therapy for Metabolic Disease”, Tsagkaraki et al 2020
CRISPR-enhanced human adipocyte ‘browning’ as cell therapy for metabolic disease
“The Burden of Rare Protein-Truncating Genetic Variants on Human Lifespan”, Liu et al 2020
The burden of rare protein-truncating genetic variants on human lifespan
“Supercentenarian and Remarkable Age Records Exhibit Patterns Indicative of Clerical Errors and Pension Fraud”, Newman 2020
“Was Jeanne Calment the Oldest Person Who Ever Lived—Or a Fraud? Some Researchers Have Cast Doubt on the Record of the Celebrated Supercentenarian”, Collins 2020
“Cryonics for All?”, Thau 2020
“Vitamin D Supplementation for Prevention of Mortality in Adults”, Bjelakovic et al 2019b
Vitamin D supplementation for prevention of mortality in adults
“A Single Combination Gene Therapy Treats Multiple Age-Related Diseases”, Davidsohn et al 2019
A single combination gene therapy treats multiple age-related diseases
“The Genetics of Human Aging”, Melzer et al 2019
“Effect of Lower Versus Higher Red Meat Intake on Cardiometabolic and Cancer Outcomes: A Systematic Review of Randomized Trials”, Zeraatkar et al 2019
“Exercise Conditioned Plasma Dampens Inflammation via Clusterin and Boosts Memory”, Miguel et al 2019
Exercise conditioned plasma dampens inflammation via clusterin and boosts memory
“A Prospective Analysis of Genetic Variants Associated With Human Lifespan”, Wright et al 2019
A Prospective Analysis of Genetic Variants Associated with Human Lifespan
“Restoration of Brain Circulation and Cellular Functions Hours Post-Mortem”, Vrselja et al 2019
Restoration of brain circulation and cellular functions hours post-mortem
“Genetically Heterogeneous Mice Exhibit a Female Survival Advantage That Is Age-Specific and Site-Specific: Results from a Large Multi-Site Study”, Cheng et al 2019
“Relaxed Selection Limits Lifespan by Increasing Mutation Load”, Cui et al 2019
Relaxed Selection Limits Lifespan by Increasing Mutation Load
“Turning Back Time With Emerging Rejuvenation Strategies”, Mahmoudi et al 2019
“An Evolutionary Perspective on Why Food Overconsumption Impairs Cognition”, Mattson 2019
An Evolutionary Perspective on Why Food Overconsumption Impairs Cognition
“A Longitudinal Big Data Approach for Precision Health”, Rose et al 2019
“What Do We Need to Know to Treat Degenerative Aging As a Medical Condition to Extend Healthy Lifespan?”, Stambler 2019
“Effects of Intermittent Fasting on Health, Aging, and Disease”, Cabo & Mattson 2019
Effects of Intermittent Fasting on Health, Aging, and Disease
“Evidence That Jeanne Calment Died in 1934-Not 1997”, Zak 2019
“Genomic Underpinnings of Lifespan Allow Prediction and Reveal Basis in Modern Risks”, Timmers et al 2018
Genomic underpinnings of lifespan allow prediction and reveal basis in modern risks
“The Protective Effect of Melatonin Against Age-Associated, Sarcopenia-Dependent Tubular Aggregate Formation, Lactate Depletion, and Mitochondrial Changes”, Sayed et al 2018
“Anti-Aging Food That Improves Markers of Health in Senior Dogs by Modulating Gut Microbiota and Metabolite Profiles”, Gebreselassie et al 2018
“Quantitative Analysis of Population-Scale Family Trees With Millions of Relatives”, Kaplanis et al 2018
Quantitative analysis of population-scale family trees with millions of relatives
“What Science Is like in North Korea: Isolated from the Rest of the World, North Korean Researchers Struggle to Balance Rigorous Scientific Work With the Demands of a Dictator”, Fiscutean 2018
“Effect of Low-Fat vs Low-Carbohydrate Diet on 12-Month Weight Loss in Overweight Adults and the Association With Genotype Pattern or Insulin SecretionThe DIETFITS Randomized Clinical Trial”, Association 2018
“Autophagy and the Cell Biology of Age-Related Disease”, Leidal et al 2018
“Early Time-Restricted Feeding Improves Insulin Sensitivity, Blood Pressure, and Oxidative Stress Even without Weight Loss in Men With Prediabetes”, Sutton et al 2018
“Selection for Long and Short Sleep Duration in Drosophila Melanogaster Reveals the Complex Genetic Network Underlying Natural Variation in Sleep”, Harbison et al 2017
“Genome-Wide Meta-Analysis Associates HLA-DQA1/DRB1 and LPA and Lifestyle Factors With Human Longevity”, Joshi et al 2017
“A Long Journey to Reproducible Results: Replicating Our Work Took Four Years and 100,000 Worms but Brought Surprising Discoveries”, Lithgow et al 2017
“Identifying Genetic Variants That Affect Viability in Large Cohorts”, Mostafavi et al 2017
Identifying genetic variants that affect viability in large cohorts
“Genetic Contribution to Two Factors of Neuroticism Is Associated With Affluence, Better Health, and Longer Life”, Hill et al 2017
“Safe Landing Strategies During a Fall: Systematic Review and Meta-Analysis”, Moon & Sosnoff 2017
Safe Landing Strategies During a Fall: Systematic Review and Meta-Analysis
“Dietary Restriction and AMPK Increase Lifespan via Mitochondrial Network and Peroxisome Remodeling”, Weir et al 2017
Dietary Restriction and AMPK Increase Lifespan via Mitochondrial Network and Peroxisome Remodeling
“2,4-Dinitrophenol, the Inferno Drug: a Netnographic Study of User Experiences in the Quest for Leanness”, McVeigh et al 2017
“Nutritional Ecology and the Evolution of Aging”, Raubenheimer et al 2016
“A Hypothesis to Explain Cancers in Confined Colonies of Naked Mole Rats”, Hochberg et al 2016
A Hypothesis to Explain Cancers in Confined Colonies of Naked Mole Rats
“Transient Rapamycin Treatment Can Increase Lifespan and Healthspan in Middle-Aged Mice”, Bitto et al 2016
Transient rapamycin treatment can increase lifespan and healthspan in middle-aged mice
“Performance of Informative Priors Skeptical of Large Treatment Effects in Clinical Trials: A Simulation Study”, Pedroza et al 2015
“The Tail End”, waitbutwhy 2015
“Consciousness without Cortex: a Hydranencephaly Family Survey”, Aleman & Merker 2014
Consciousness without cortex: a hydranencephaly family survey
“Vitamin D and Vitamin D Analogues for Preventing Fractures in Post-Menopausal Women and Older Men”, Avenell et al 2014
Vitamin D and vitamin D analogues for preventing fractures in post-menopausal women and older men
“Vitamin D Supplementation for Prevention of Cancer in Adults (Review)”, G et al 2014
Vitamin D supplementation for prevention of cancer in adults (Review)
“A Review of Creatine Supplementation in Age-Related Diseases: More Than a Supplement for Athletes”, Smith et al 2014
A review of creatine supplementation in age-related diseases: more than a supplement for athletes
“Who By Very Slow Decay”, Alexander 2013
“Reproductive Capability Is Associated With Lifespan and Cause of Death in Companion Dogs”, Hoffman et al 2013
Reproductive Capability Is Associated with Lifespan and Cause of Death in Companion Dogs
“Multivitamin-Multimineral Supplementation and Mortality: a Meta-Analysis of Randomized Controlled Trials”, Macpherson et al 2013
“The Caenorhabditis Elegans Lifespan Machine”, Stroustrup et al 2013
“‘Life in a Germ-Free World’: Isolating Life from the Laboratory Animal to the Bubble Boy § Germ-Free Benefits”, Kirk 2012 (page 7)
“Terminal Dedifferentiation of Cognitive Abilities”, Wilson et al 2012
“A Brief History of Cross-Species Organ Transplantation”, Cooper 2012
“Sitting Time and All-Cause Mortality Risk in 222,497 Australian Adults”, Ploeg 2012
Sitting Time and All-Cause Mortality Risk in 222,497 Australian Adults
“Melatonin in Aging and Disease -Multiple Consequences of Reduced Secretion, Options and Limits of Treatment”, Hardeland 2012
“Happy Aged People Are All Alike, While Every Unhappy Aged Person Is Unhappy in Its Own Way”, Tumminello et al 2011
Happy Aged People Are All Alike, While Every Unhappy Aged Person Is Unhappy in Its Own Way
“Vitamin D and Cardiovascular Outcomes: A Systematic Review and Meta-Analysis”, Elamin et al 2011
Vitamin D and Cardiovascular Outcomes: A Systematic Review and Meta-Analysis
“Oral Anti-Diabetic Drugs for the Prevention of Type 2 Diabetes”, Phung 2011
Oral anti-diabetic drugs for the prevention of type 2 diabetes
“Intelligence in Youth and All-Cause-Mortality: Systematic Review With Meta-Analysis”, Calvin et al 2011
Intelligence in youth and all-cause-mortality: systematic review with meta-analysis
“Compression of Morbidity 1980–2011: a Focused Review of Paradigms and Progress”, Fries et al 2011
Compression of morbidity 1980–2011: a focused review of paradigms and progress
“A Genome-Wide Association Study of Aging”, Walter et al 2011
“Association of Circulating Visfatin Concentrations With Insulin Resistance and Low-Grade Inflammation After Dietary Energy Restriction in Spanish Obese Non-Diabetic Women: Role of Body Composition Changes”, Agueda et al 2010
“Japan, Checking on Its Oldest, Finds Many Gone”
“Demographic Consequences of Defeating Aging”, Gavrilov & Gavrilova 2010
“Biodemography of Human Aging”, Vaupel 2010
“Typologies of Extreme Longevity Myths”, Young et al 2010
“Ageing and Its Implications”, Jayanthi et al 2010
“Plasma Vitamin D and Mortality in Older Men: a Community-Based Prospective Cohort Study”, Michaëlsson 2010
Plasma vitamin D and mortality in older men: a community-based prospective cohort study
“A Weak Link in Metabolism: the Metabolic Capacity for Glycine Biosynthesis Does Not Satisfy the Need for Collagen Synthesis”, Meléndez-Hevia et al 2009
“Good Semen Quality and Life Expectancy: A Cohort Study of 43,277 Men”, Jensen et al 2009
Good Semen Quality and Life Expectancy: A Cohort Study of 43,277 Men
“Factors Contributing to the Facial Aging of Identical Twins”, Guyuron et al 2009
“Use of Supplements of Multivitamins, Vitamin C, and Vitamin E in Relation to Mortality”, Pocobelli 2009
Use of Supplements of Multivitamins, Vitamin C, and Vitamin E in Relation to Mortality
“Vitamin D and Aging”, Tuohimaa 2009 (page 2)
“Ageing Populations: the Challenges Ahead”, Christensen et al 2009
“Effects of Calcium and Vitamin D Supplementation on Hip Bone Mineral Density and Calcium-Related Analytes in Elderly Ambulatory Australian Women: A Five-Year Randomized Controlled Trial”, Zhu et al 2008
“Prolonged Survival to Adulthood of a [22yo] Individual With Hydranencephaly”, Bae et al 2008
Prolonged survival to adulthood of a [22yo] individual with hydranencephaly
“Influence of Environmental Factors on Facial Aging”, Rexbye et al 2006
“Understanding the Odd Science of Aging”, Kirkwood 2005
“A Randomized, Controlled Trial of Quadriceps Resistance Exercise and Vitamin D in Frail Older People: The Frailty Interventions Trial in Elderly Subjects (FITNESS)”, Latham et al 2003
“The Reserve-Capacity Hypothesis: Evolutionary Origins and Modern Implications of the Trade-Off between Tumor-Suppression and Tissue-Repair”, Weinstein & Ciszek 2002
“Book Review: Validation of Exceptional Longevity, by Bernard Jeune and James W. Vaupel”, Gavrilov & Gavrilova 2000
Book review: Validation of Exceptional Longevity, by Bernard Jeune and James W. Vaupel
“Prevention of Femoral and Lumbar Bone Loss With Hormone Replacement Therapy and Vitamin D3 in Early Postmenopausal Women: A Population-Based 5–Year Randomized Trial”, Komulainen et al 1999
“A C. Elegans Mutant That Lives Twice As Long As Wild Type”, Kenyon et al 1993
Pritikin: The Man Who Healed America's Heart, Monte & Pritikin 1988
Disguised, Moore & Conn 1985
“Eternal Youth Kills Ants”, Times 1982
“What Difference Would It Make If Cancer Were Eradicated? An Examination of the Taeuber Paradox”, Keyfitz 1977
What Difference Would It Make if Cancer Were Eradicated? An Examination of the Taeuber Paradox
“Calmly We Walk through This April’s Day”, Schwartz 1967
“Aging in Germ-Free Mice: Life Tables and Lesions Observed at Natural Death”, Gordon et al 1966
Aging in Germ-free Mice: Life Tables and Lesions Observed at Natural Death
“Influence Of The Normal Flora On Mucosal Morphology And Cellular Renewal In The Ileum. A Comparison Of Germ-Free And Conventional Mice”, Abrams et al 1963
“Three Jubilees: L. Tolstoy, Hen. Ibsen, N. Fedorov”
“The Machine Man of Ardathia”, Flagg 1927
“Spring and Fall”, Hopkins 1880
“The Value of Health and Longevity”
“The Old Fools”, Larkin 2026
“Association of Neurocognitive and Physical Function With Gait Speed in Midlife Neurology”
Association of Neurocognitive and Physical Function With Gait Speed in Midlife Neurology
“Nintil”, Ricon 2026
“Low-Dose Lithium Uptake Promotes Longevity in Humans and Metazoans”
Low-dose lithium uptake promotes longevity in humans and metazoans
“Eulogy to the Obits”, Balwit 2026
“Your Book Review: The Family That Couldn’t Sleep”
“More Than 230,000 Japanese Centenarians ‘Missing’”
“Computational Analysis of Lifespan Experiment Reproducibility”
Computational Analysis of Lifespan Experiment Reproducibility
“Core Pathways of Aging”
View External Link:
https://www.lesswrong.com/posts/ui6mDLdqXkaXiDMJ5/core-pathways-of-aging
“Shinya Yamanaka – Biographical”
“James F. Fries, Who Studied the Good Life and How to Live It, Dies at 83”
James F. Fries, Who Studied the Good Life and How to Live It, Dies at 83
“Dietary Magnesium Intake Is Inversely Associated With Mortality in Adults at High Cardiovascular Disease Risk”
SkiFree, Munroe 2026
View External Link:
“J’accuse…! Why Jeanne Calment’s 122-Year Old Longevity Record May Be Fake”
J’accuse…! Why Jeanne Calment’s 122-year old longevity record may be fake
“More Evidence for Jeanne Calment’s Identity Theft Hypothesis”
More evidence for Jeanne Calment’s identity theft hypothesis
Sort By Magic
Annotations sorted by machine learning into inferred 'tags'. This provides an alternative way to browse: instead of by date order, one can browse in topic order. The 'sorted' list has been automatically clustered into multiple sections & auto-labeled for easier browsing.
Beginning with the newest annotation, it uses the embedding of each annotation to attempt to create a list of nearest-neighbor annotations, creating a progression of topics. For more details, see the link.
renewal
cellular-turnover
fat-loss
longevity-investments
brain-repair
extreme-longevity
healthspan-advancement
Wikipedia (8)
Miscellaneous
/doc/longevity/2020-bischoffferrari-table1-summaryresults.png/doc/longevity/2016-hochberg-figure1-nakedmoleratsurvivalcurvesinwildvscaptivity.jpg/doc/longevity/2013-grg-oldestperson-successiveintervals.png/doc/longevity/2009-guyuron-figure3-effectofsmokingonfacialaginginpairsofidenticaltwins.jpg/doc/longevity/2009-guyuron-figure4-smokingtwinlooksolderthanlesssmokingidenticaltwin.jpg/doc/longevity/2009-guyuron-figure5-smokingtwinlooksolderthanlesssmokingidenticaltwinmales.png/doc/longevity/2009-guyuron-figure6-identicaltwinsdiscordantinsunexposureandbmiagedifference.jpg/doc/philosophy/ethics/2004-04-12-sage-leonkass-agelessbodiesandhappysoulsinterview.html/doc/longevity/1977-friedman-proteincrosslinkingnutritionalandmedicalconsequences.pdf/doc/longevity/gwern-gompertzcurve-longevity-calmentcasestudy.png/doc/longevity/gwern-gompertzcurve-longevity-calmentcasestudy-unsorted.pnghttps://link.springer.com/article/10.1007/s11357-023-00818-1https://thelampmagazine.com/issues/issue-17/shadow-on-the-sunhttps://trevorklee.substack.com/p/is-longevity-a-choice-how-about-obesityhttps://vmc.vet.osu.edu/cto/clinical-trials/rapamycin-cats-chronic-kidney-diseasehttps://worksinprogress.co/issue/the-future-of-kidney-treatment/https://www.demographic-research.org/volumes/vol40/29/40-29.pdfhttps://www.demographic-research.org/volumes/vol49/27/49-27.pdfhttps://www.frontiersin.org/journals/endocrinology/articles/10.3389/fendo.2021.722187/fullhttps://www.sciencedirect.com/science/article/pii/S0047637419300181https://www.statnews.com/2022/08/09/anti-aging-projects-funding-much-discussed-trial-overlooked/https://www.thelancet.com/journals/lanhl/article/PIIS2666-7568(23)00189-7/fulltexthttps://www.theonion.com/world-death-rate-holding-steady-at-100-percent-1819564171https://www.wired.com/story/patricia-moore-sacrificed-youth-to-get-tech-bros-to-grow-up/
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Bibliography
https://academic.oup.com/eurjpc/advance-article/doi/10.1093/eurjpc/zwaf267/8128214: “Cardiorespiratory Fitness in Adolescence and Premature Mortality: Widespread Bias Identified Using Negative Control Outcomes and Sibling Comparisons”,https://www.nytimes.com/2025/03/21/technology/bryan-johnson-blueprint-confidentiality-agreements.html: “How Bryan Johnson, Who Wants to Live Forever, Sought Control via Confidentiality Agreements: That Control Is Starting to Fray As Johnson, a Longevity Guru Known for Performing Experiments on His Body, Faces Backlash”,https://papers.ssrn.com/sol3/papers.cfm?abstract_id=4916412: “(Inaccurate) Beliefs about Skill Decay”,https://www.technologyreview.com/2023/03/08/1069523/sam-altman-investment-180-million-retro-biosciences-longevity-death/: “Sam Altman Invested $180 Million into a Company Trying to Delay Death: Can Anti-Aging Breakthroughs Add 10 Healthy Years to the Human Life Span? The CEO of OpenAI Is Paying to Find Out”,https://onlinelibrary.wiley.com/doi/10.1111/acel.13724: “Lifespan Benefits for the Combination of Rapamycin plus Acarbose and for Captopril in Genetically Heterogeneous Mice”,https://link.springer.com/article/10.1007/s11357-022-00645-w: “Old Plasma Dilution Reduces Human Biological Age: a Clinical Study”,https://www.nature.com/articles/s43587-021-00159-8: “Mendelian Randomization of Genetically Independent Aging Phenotypes Identifies LPA and VCAM1 As Biological Targets for Human Aging”,https://journals.plos.org/plosgenetics/article?id=10.1371/journal.pgen.1007098: “Selection for Long and Short Sleep Duration in Drosophila Melanogaster Reveals the Complex Genetic Network Underlying Natural Variation in Sleep”,2014-aleman.pdf: “Consciousness without Cortex: a Hydranencephaly Family Survey”,2012-kirk.pdf#page=7: “‘Life in a Germ-Free World’: Isolating Life from the Laboratory Animal to the Bubble Boy § Germ-Free Benefits”,https://www.sciencedirect.com/science/article/pii/S0092867405001017: “Understanding the Odd Science of Aging”,1966-gordon.pdf: “Aging in Germ-Free Mice: Life Tables and Lesions Observed at Natural Death”,