- See Also
- Gwern
-
Links
- “Human Deleterious Mutation Rate Implies High Fitness Variance, With Declining Mean Fitness Compensated by Rarer Beneficial Mutations of Larger Effect”, Matheson et al 2023
- “Husband’s Income, Wife’s Income, and Number of Biological Children in the U.S.”, Hopcroft 2022
- “Contemporary Selection Pressures in Modern Societies? Which Factors Best Explain Variance in Human Reproduction and Mating?”, Fieder & Huber 2022
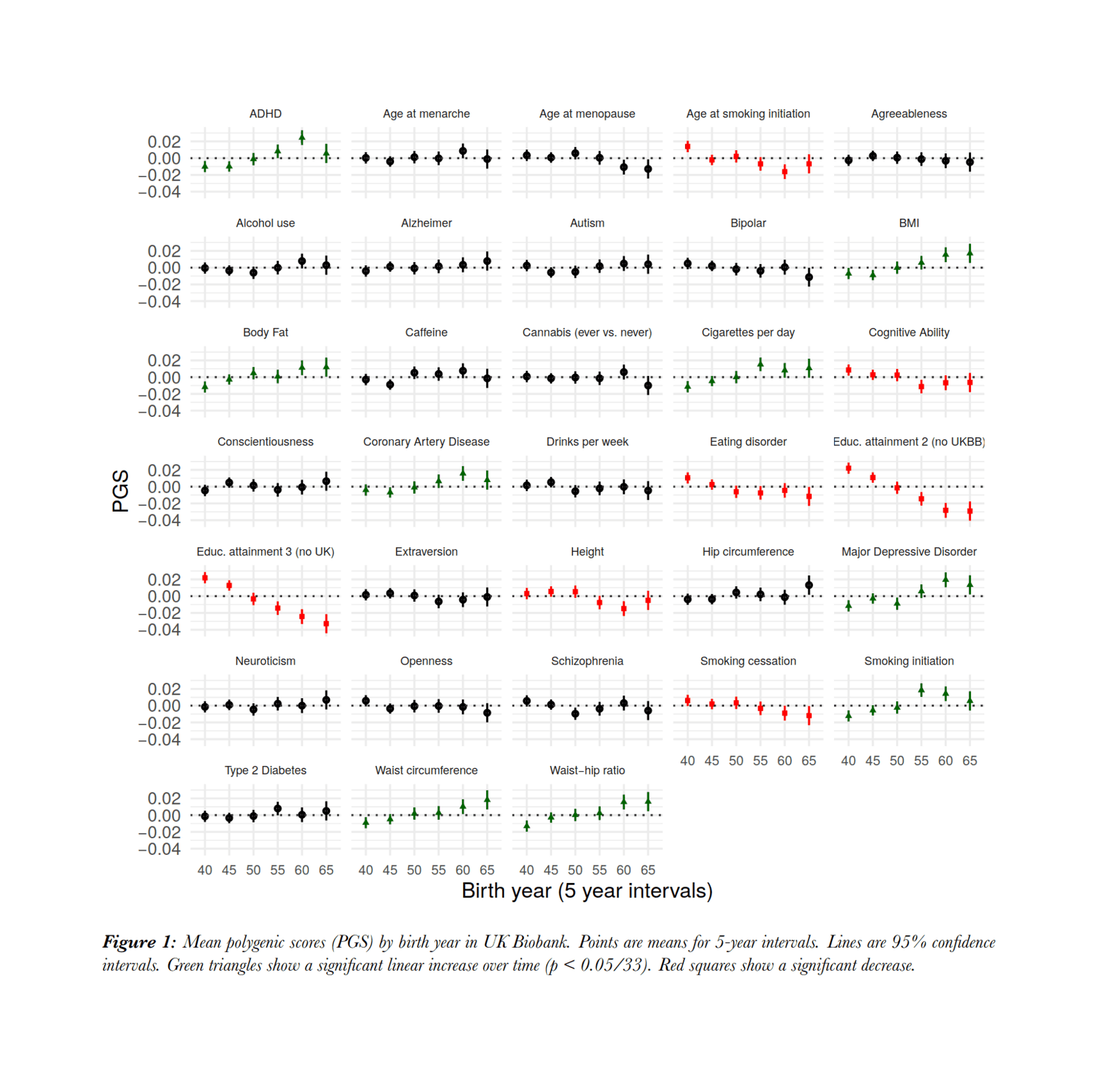
- “A Selection Pressure Landscape for 870 Human Polygenic Traits”, Song et al 2021
- “The Children of the Missed Pill”, Rau et al 2021
- “High Income Men Have High Value As Long-Term Mates in the U.S.: Personal Income and the Probability of Marriage, Divorce, and Childbearing in the U.S.”, Hopcroft 2021
- “The Secular Trend of Intelligence Test Scores in the Present Century: The Danish Experience”, Hegelund et al 2021
- “Identification of 370 Genetic Loci for Age at First Sex and Birth Linked to Externalizing Behavior”, Mills et al 2021
- “Natural Selection in Contemporary Humans Is Linked to Income and Substitution Effects”, Hugh-Jones & Abdellaoui 2021
- “The Micro-Evidence for the Malthusian System: France, 1670–1840”, Cummins 2020
- “Germline Mutation Rates in Young Adults Predict Longevity and Reproductive Lifespan”, Cawthon et al 2020
- “Genome-Wide Analysis Identifies Genetic Effects on Reproductive Success and Ongoing Natural Selection at the FADS Locus”, Mathieson et al 2020
- “Sociology, Genetics, and the Coming of Age of Sociogenomics”, Mills & Tropf 2020
- “Associations of Autozygosity With a Broad Range of Human Phenotypes”, Clark et al 2019
- “Throwing the Baby Out With the Bath Water: Could Widespread Neutering of Companion Dogs Cause Problems at a Population Level?”, Dawson et al 2019
- “The Genetics of Human Fertility”, Kim & Lee 2019
- “How Intelligence Affects Fertility 30 Years On: Retherford and Sewell Revisited—With Polygenic Scores and Numbers of Grandchildren”, Menie et al 2019b
- “A Systematic Review of the State of Literature Relating Parental General Cognitive Ability and Number of Offspring”, Reeve et al 2018
- “Evidence of a Nonadaptive Buildup of Mutational Load in Human Populations over the past 40,000 Years”, Aris-Brosou 2018
- “The Transition to Modernity and Chronic Disease: Mismatch and Natural Selection”, Corbett et al 2018
- “The Contemporary Evolution of Fitness”, Hendry et al 2018
- “Signatures of Negative Selection in the Genetic Architecture of Human Complex Traits”, Zeng et al 2018
- “Sexual Dimorphism in the Genetic Influence on Human Childlessness”, Verweij et al 2017
- “The Genomic Health of Ancient Hominins”, Berens et al 2017
- “Holocene Selection for Variants Associated With Cognitive Ability: Comparing Ancient and Modern Genomes”, Menie et al 2017
- “Selection against Variants in the Genome Associated With Educational Attainment”, Kong et al 2017
- “Evidence of Directional and Stabilizing Selection in Contemporary Humans”
- “Genome-Wide Analysis Identifies 12 Loci Influencing Human Reproductive Behavior”, Barban et al 2016
- “How Cognitive Genetic Factors Influence Fertility Outcomes: A Mediational SEM Analysis”, Menie et al 2016
- “Evidence of Dysgenic Fertility in China”, Wang et al 2016c
- “Assortative Mating and Differential Fertility by Phenotype and Genotype across the 20th Century”, Conley et al 2016
- “Mega-Analysis of 31,396 Individuals from 6 Countries Uncovers Strong Gene-Environment Interaction for Human Fertility”, Tropf et al 2016
- “Mortality Selection in a Genetic Sample and Implications for Association Studies”, Domingue et al 2016
- “Older Fathers’ Children Have Lower Evolutionary Fitness across Four Centuries and in Four Populations”, Arslan et al 2016
- “Determinants of Online Sperm Donor Success: How Women Choose”, Whyte & Torgler 2015
- “The Biodemography of Fertility: A Review and Future Research Frontiers”, Mills & Tropf 2015
- “Infertility Etiologies Are Genetically and Clinically Linked With Other Diseases in Single Meta-Diseases”, Tarín et al 2015
- “Fecundity of Patients With Schizophrenia, Autism, Bipolar Disorder, Depression, Anorexia Nervosa, or Substance Abuse vs Their Unaffected Siblings”, Power et al 2013
- “Physical Attractiveness and Reproductive Success in Humans: Evidence from the Late 20th Century United States”, Jokela 2009
- “Secular Declines in Cognitive Test Scores: A Reversal of the Flynn Effect”, Teasdale & Owen 2008
- “Feral Cats: Their Role in the Population Dynamics of Felis Catus”, Bradshaw et al 1999
- “DYSGENICS: Genetic Deterioration in Modern Populations”, Writer 1996
- “New Methodology to Reduce the Environment-Heredity Uncertainty About Dysgenics”, Shockley 1970
- “Interplay Between Social Ecology and Physiology, Genetics and Population Dynamics of Mice”, Kessler 1966
- “You and Some 'Cavemen' Get a Genetic Checkup”
- “Why the Red Delicious No Longer Is”
- Sort By Magic
- Miscellaneous
- Link Bibliography
See Also
Gwern
“Statistical Notes”, Gwern 2014
“Cat Psychology & Domestication: Are We Good Owners?”, Gwern 2018
Links
“Human Deleterious Mutation Rate Implies High Fitness Variance, With Declining Mean Fitness Compensated by Rarer Beneficial Mutations of Larger Effect”, Matheson et al 2023
“Husband’s Income, Wife’s Income, and Number of Biological Children in the U.S.”, Hopcroft 2022
Husband’s income, wife’s income, and number of biological children in the U.S.
“Contemporary Selection Pressures in Modern Societies? Which Factors Best Explain Variance in Human Reproduction and Mating?”, Fieder & Huber 2022
“A Selection Pressure Landscape for 870 Human Polygenic Traits”, Song et al 2021
A selection pressure landscape for 870 human polygenic traits
“The Children of the Missed Pill”, Rau et al 2021
“High Income Men Have High Value As Long-Term Mates in the U.S.: Personal Income and the Probability of Marriage, Divorce, and Childbearing in the U.S.”, Hopcroft 2021
“The Secular Trend of Intelligence Test Scores in the Present Century: The Danish Experience”, Hegelund et al 2021
The secular trend of intelligence test scores in the present century: The Danish experience
“Identification of 370 Genetic Loci for Age at First Sex and Birth Linked to Externalizing Behavior”, Mills et al 2021
Identification of 370 genetic loci for age at first sex and birth linked to externalizing behavior
“Natural Selection in Contemporary Humans Is Linked to Income and Substitution Effects”, Hugh-Jones & Abdellaoui 2021
Natural Selection in Contemporary Humans is Linked to Income and Substitution Effects
“The Micro-Evidence for the Malthusian System: France, 1670–1840”, Cummins 2020
The micro-evidence for the Malthusian system: France, 1670–1840
“Germline Mutation Rates in Young Adults Predict Longevity and Reproductive Lifespan”, Cawthon et al 2020
Germline mutation rates in young adults predict longevity and reproductive lifespan
“Genome-Wide Analysis Identifies Genetic Effects on Reproductive Success and Ongoing Natural Selection at the FADS Locus”, Mathieson et al 2020
“Sociology, Genetics, and the Coming of Age of Sociogenomics”, Mills & Tropf 2020
“Associations of Autozygosity With a Broad Range of Human Phenotypes”, Clark et al 2019
Associations of autozygosity with a broad range of human phenotypes
“Throwing the Baby Out With the Bath Water: Could Widespread Neutering of Companion Dogs Cause Problems at a Population Level?”, Dawson et al 2019
“The Genetics of Human Fertility”, Kim & Lee 2019
“How Intelligence Affects Fertility 30 Years On: Retherford and Sewell Revisited—With Polygenic Scores and Numbers of Grandchildren”, Menie et al 2019b
“A Systematic Review of the State of Literature Relating Parental General Cognitive Ability and Number of Offspring”, Reeve et al 2018
“Evidence of a Nonadaptive Buildup of Mutational Load in Human Populations over the past 40,000 Years”, Aris-Brosou 2018
Evidence of a nonadaptive buildup of mutational load in human populations over the past 40,000 years
“The Transition to Modernity and Chronic Disease: Mismatch and Natural Selection”, Corbett et al 2018
The transition to modernity and chronic disease: mismatch and natural selection:
“The Contemporary Evolution of Fitness”, Hendry et al 2018
“Signatures of Negative Selection in the Genetic Architecture of Human Complex Traits”, Zeng et al 2018
Signatures of negative selection in the genetic architecture of human complex traits:
“Sexual Dimorphism in the Genetic Influence on Human Childlessness”, Verweij et al 2017
Sexual dimorphism in the genetic influence on human childlessness
“The Genomic Health of Ancient Hominins”, Berens et al 2017
“Holocene Selection for Variants Associated With Cognitive Ability: Comparing Ancient and Modern Genomes”, Menie et al 2017
“Selection against Variants in the Genome Associated With Educational Attainment”, Kong et al 2017
Selection against variants in the genome associated with educational attainment
“Evidence of Directional and Stabilizing Selection in Contemporary Humans”
Evidence of directional and stabilizing selection in contemporary humans
“Genome-Wide Analysis Identifies 12 Loci Influencing Human Reproductive Behavior”, Barban et al 2016
Genome-wide analysis identifies 12 loci influencing human reproductive behavior
“How Cognitive Genetic Factors Influence Fertility Outcomes: A Mediational SEM Analysis”, Menie et al 2016
How cognitive genetic factors influence fertility outcomes: A mediational SEM analysis
“Evidence of Dysgenic Fertility in China”, Wang et al 2016c
“Assortative Mating and Differential Fertility by Phenotype and Genotype across the 20th Century”, Conley et al 2016
Assortative mating and differential fertility by phenotype and genotype across the 20th century
“Mega-Analysis of 31,396 Individuals from 6 Countries Uncovers Strong Gene-Environment Interaction for Human Fertility”, Tropf et al 2016
“Mortality Selection in a Genetic Sample and Implications for Association Studies”, Domingue et al 2016
Mortality Selection in a Genetic Sample and Implications for Association Studies
“Older Fathers’ Children Have Lower Evolutionary Fitness across Four Centuries and in Four Populations”, Arslan et al 2016
“Determinants of Online Sperm Donor Success: How Women Choose”, Whyte & Torgler 2015
Determinants of online sperm donor success: how women choose
“The Biodemography of Fertility: A Review and Future Research Frontiers”, Mills & Tropf 2015
The Biodemography of Fertility: A Review and Future Research Frontiers
“Infertility Etiologies Are Genetically and Clinically Linked With Other Diseases in Single Meta-Diseases”, Tarín et al 2015
“Fecundity of Patients With Schizophrenia, Autism, Bipolar Disorder, Depression, Anorexia Nervosa, or Substance Abuse vs Their Unaffected Siblings”, Power et al 2013
“Physical Attractiveness and Reproductive Success in Humans: Evidence from the Late 20th Century United States”, Jokela 2009
“Secular Declines in Cognitive Test Scores: A Reversal of the Flynn Effect”, Teasdale & Owen 2008
Secular declines in cognitive test scores: A reversal of the Flynn Effect
“Feral Cats: Their Role in the Population Dynamics of Felis Catus”, Bradshaw et al 1999
Feral cats: their role in the population dynamics of Felis catus
“DYSGENICS: Genetic Deterioration in Modern Populations”, Writer 1996
“New Methodology to Reduce the Environment-Heredity Uncertainty About Dysgenics”, Shockley 1970
New Methodology to Reduce the Environment-Heredity Uncertainty About Dysgenics:
“Interplay Between Social Ecology and Physiology, Genetics and Population Dynamics of Mice”, Kessler 1966
Interplay Between Social Ecology and Physiology, Genetics and Population Dynamics of Mice
“You and Some 'Cavemen' Get a Genetic Checkup”
“Why the Red Delicious No Longer Is”
Sort By Magic
Annotations sorted by machine learning into inferred 'tags'. This provides an alternative way to browse: instead of by date order, one can browse in topic order. The 'sorted' list has been automatically clustered into multiple sections & auto-labeled for easier browsing.
Beginning with the newest annotation, it uses the embedding of each annotation to attempt to create a list of nearest-neighbor annotations, creating a progression of topics. For more details, see the link.
intelligence-trend
reproductive-genetics
fertility-genetics
Miscellaneous
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Link Bibliography
-
2022-hopcroft.pdf: “Husband’s Income, Wife’s Income, and Number of Biological Children in the U.S.”, Rosemary L. Hopcroft -
2021-song.pdf: “A Selection Pressure Landscape for 870 Human Polygenic Traits”, Weichen Song, Yueqi Shi, Weidi Wang, Weihao Pan, Wei Qian, Shunying Yu, Min Zhao, Guan Ning Lin -
https://ideas.repec.org/p/uea/ueaeco/2021-02.html: “Natural Selection in Contemporary Humans Is Linked to Income and Substitution Effects”, David Hugh-Jones, Abdel Abdellaoui -
2019-woodley-2.pdf: “How Intelligence Affects Fertility 30 Years On: Retherford and Sewell Revisited—With Polygenic Scores and Numbers of Grandchildren”, Michael A. Woodley of Menie, Heiner Rindermann, Jonatan Pallesen, Matthew A. Sarraf