- See Also
-
Links
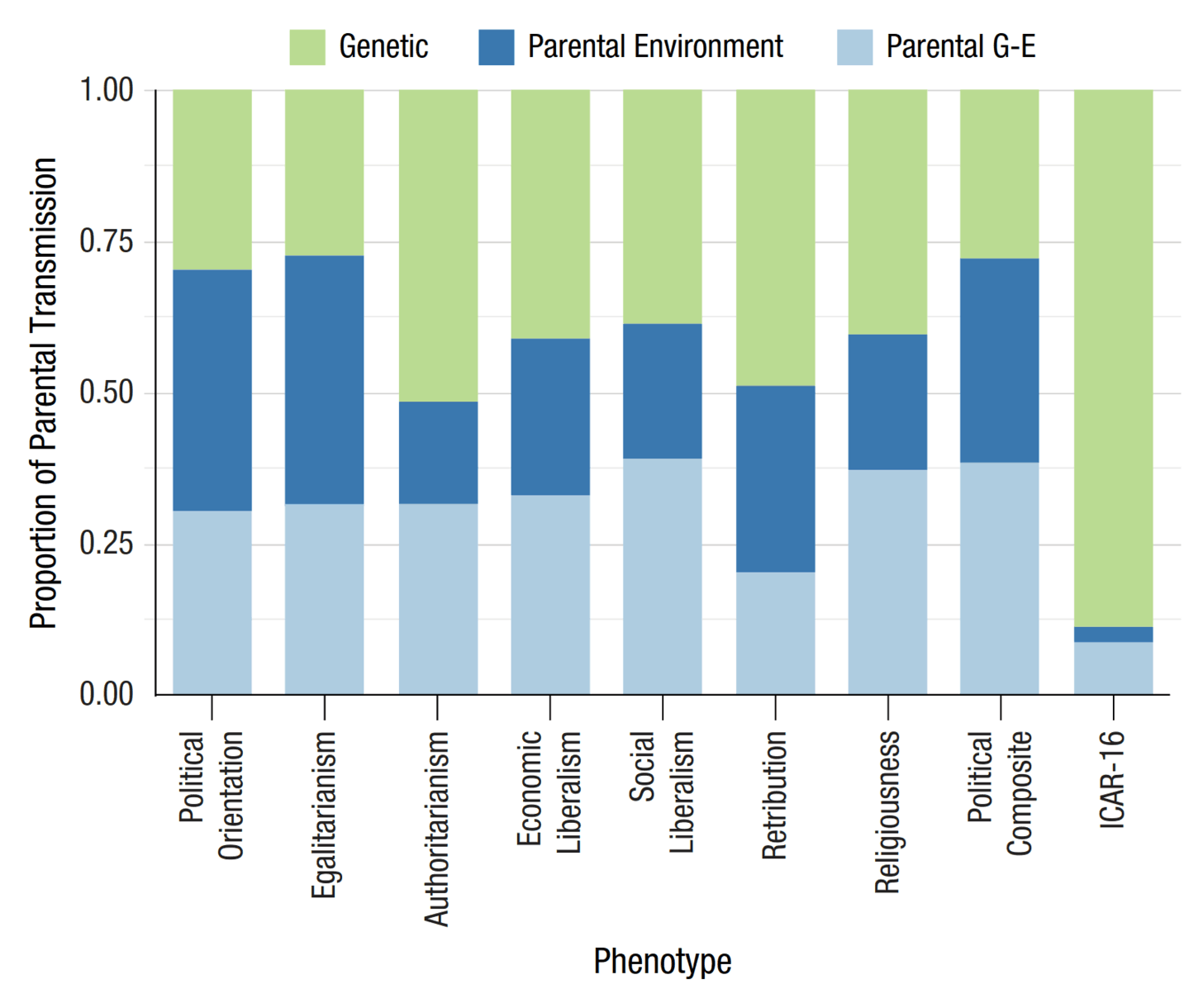
- “Predicting Political Beliefs With Polygenic Scores for Cognitive Performance and Educational Attainment”, Edwards et al 2024
- “The Garden of Forking Paths; An Evaluation of Joseph’s ‘A Reevaluation of the 1990 Minnesota Study of Twins Reared Apart IQ Study’”, Bouchard 2023
- “Personality Traits, Mental Abilities and Other Individual Differences: Monozygotic Female Twins Raised Apart in South Korea and the United States”, Segal & Hur 2022
- “Parent Contributions to the Development of Political Attitudes in Adoptive and Biological Families”, Willoughby et al 2021b
- “Genetic and Environmental Contributions to IQ in Adoptive and Biological Families With 30-Year-Old Offspring”, Willoughby et al 2021
- “Heritability × SES Interaction for IQ: Is It Present in US Adoption Studies?”, Loehlin et al 2021
- “Does Biology Drive Child Penalties? Evidence from Biological and Adoptive Families”, Kleven et al 2021
- “Parental Income and Mental Disorders in Children and Adolescents: Prospective Register-Based Study”, Kinge et al 2021
- “Why Do Wealthy Parents Have Wealthy Children?”, Fagereng et al 2021
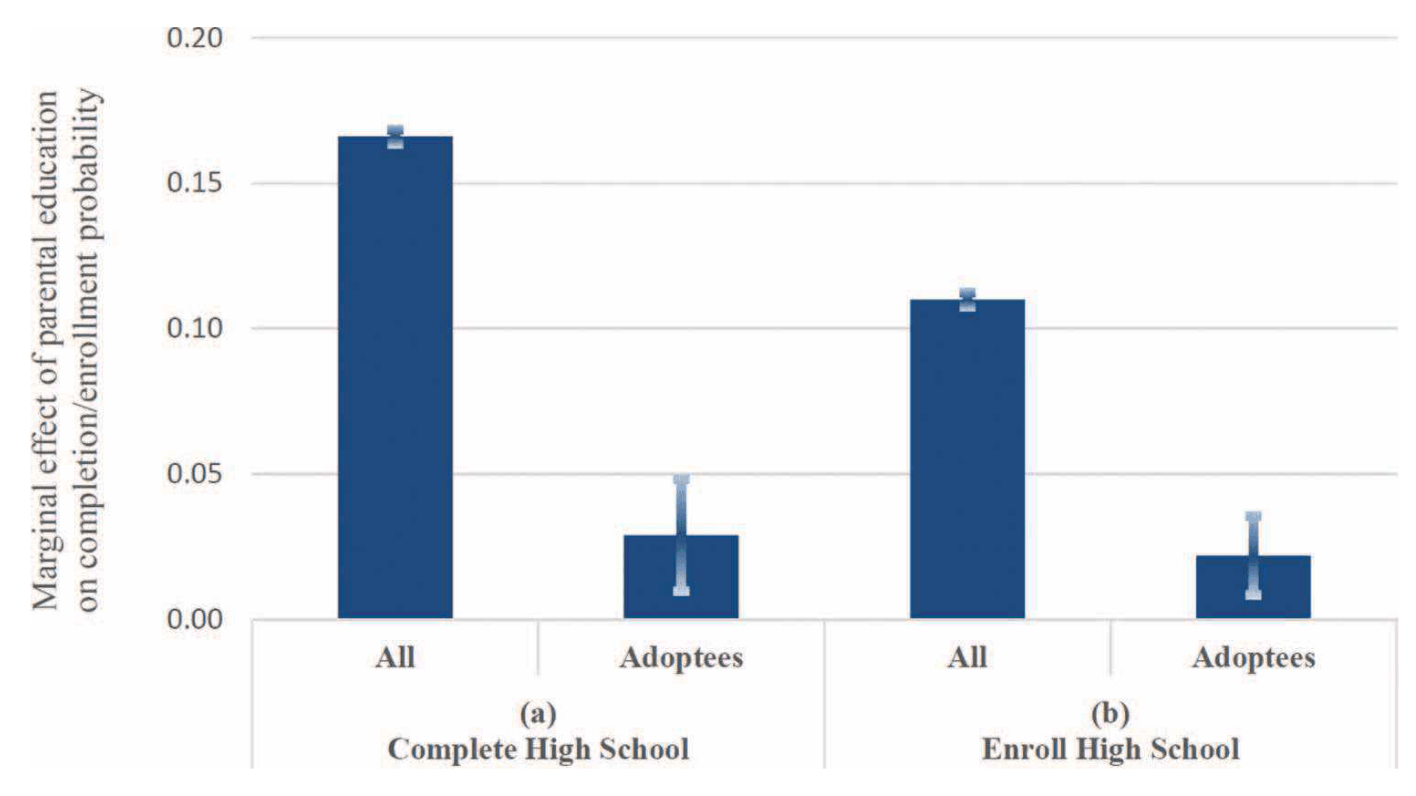
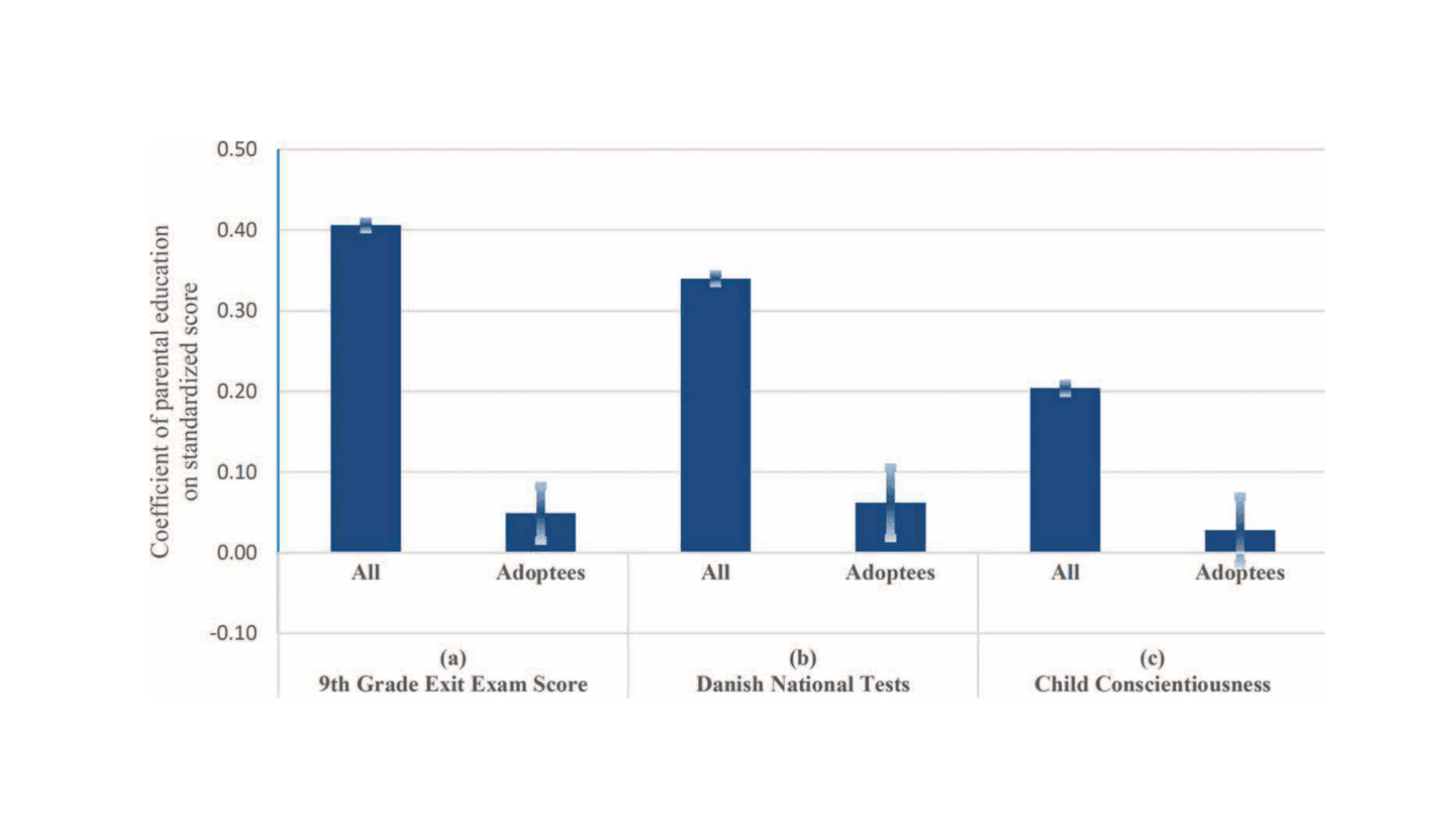
- “Does Parental Education Influence Child Educational Outcomes? A Developmental Analysis in a Full-Population Sample and Adoptee Design”, Ludeke et al 2021
- “The Role of the Shared Environment in College Attainment: An Adoption Study”, Anderson et al 2020b
- “The Intergenerational Transmission of Early Educational Advantages: New Results Based on an Adoption Design”, Halpern-Manners et al 2020
- “Childhood Adoption and Mental Health in Adulthood: The Role of Gene-Environment Correlations and Interactions in the UK Biobank”, Lehto et al 2019
- “The Association Between Genetic Predisposition and Parental Socialization: An Examination of Gene-Environment Correlations Using an Adoption-Based Design”, Knoblach et al 2019
- “Fullerton Virtual Twin Project: Overview and 2019 Update”, Segal & Niculae 2019
- “Genetics, the Rearing Environment, and the Intergenerational Transmission of Divorce: A Swedish National Adoption Study”, Salvatore et al 2018
- “Childhood Social Class and Cognitive Aging in the Swedish Adoption/Twin Study of Aging”, Ericsson et al 2017
- “Cross-Generational Transmission from Drug Abuse in Parents to Attention-Deficit/hyperactivity Disorder in Children”, Kendler et al 2016
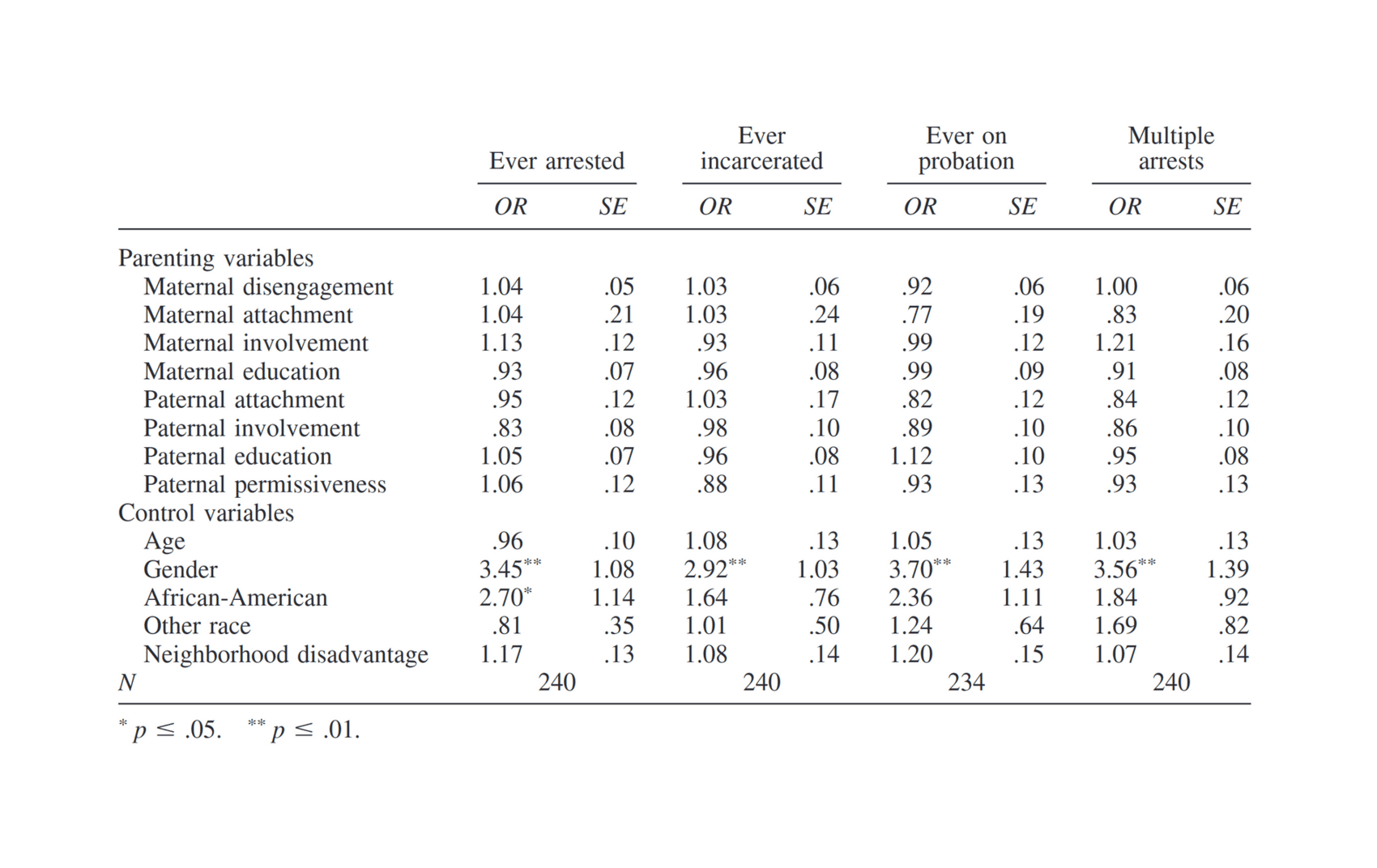
- “The Role of Parenting in the Prediction of Criminal Involvement: Findings from a Nationally Representative Sample of Youth and a Sample of Adopted Youth”, Beaver et al 2015
- “Are Adoption Gains on the G Factor? A Meta-Analysis”, Nijenhuis et al 2015
- “The Heritability of Alcohol Use Disorders: a Meta-Analysis of Twin and Adoption Studies”, Verhulst et al 2015
- “Family Environment and the Malleability of Cognitive Ability: a Swedish National Home-Reared and Adopted-Away Cosibling Control Study”, Kendler et al 2015
- “The Relationship between Parental Depressive Symptoms and Offspring Psychopathology: Evidence from a Children-Of-Twins Study and an Adoption Study”, McAdams et al 2015
- “Born in Korea-Adopted Apart: Behavioral Development of Monozygotic Twins Raised in the United States and France”, Segal & Cortez 2014
- “Children of Twins Design”, D’Onofrio 2014
- “A Closer Look at the Role of Parenting-Related Influences on Verbal Intelligence over the Life Course: Results from an Adoption-Based Research Design”, Beaver et al 2014
- “‘Mischievous Responders’ Confound Research On Teens”, Kamenetz 2014
- “Inaccurate Estimation of Disparities Due to Mischievous Responders”, Robinson-Cimpian 2014
- “Continuity of Genetic and Environmental Influences on Cognition across the Life Span: A Meta-Analysis of Longitudinal Twin and Adoption Studies”, Tucker-Drob & Briley 2014
- “Quantitative Genetics in the Postmodern Family of the Donor Sibling Registry”, Lee 2013b
- “The Lost Study: A 1998 Adoption Study of Personality That Found No Genetic Relationship between Birth-Parents and Their 240 Adopted-Away Biological Offspring”, Joseph 2013
- “Intelligence: New Findings and Theoretical Developments”, Nisbett et al 2012
- “Genetic and Familial Environmental Influences on the Risk for Drug Abuse: a National Swedish Adoption Study”, Kendler et al 2012
- “The Causal Effect of Parents' Schooling on Children’s Schooling: A Comparison of Estimation Methods”, Holmlund et al 2011
- “Why Are Children in the Same Family so Different from One Another?”, Plomin & Daniels 2011
- “Human Aggression across the Lifespan: Genetic Propensities and Environmental Moderators”, Tuvblad & Baker 2011
- “Genetic and Environmental Influences on Adult Life Outcomes: Evidence from the Texas Adoption Project”, Loehlin et al 2007
- “How Large Are the Effects from Changes in Family Environment? A Study of Korean American Adoptees”, Sacerdote 2007
- “Proceeding From Observed Correlation to Causal Inference: The Use of Natural Experiments”, Rutter 2007
- “The Mechanism of Emergenesis”, Lykken 2006
- “Biology-Environment Interaction and Evocative Biology-Environment Correlation: Contributions of Harsh Discipline and Parental Psychopathology to Problem Adolescent Behaviors”, Riggins-Caspers et al 2003
- “The Nature and Nurture of Economic Outcomes”, Sacerdote 2002
- “Implications of the Restricted Range of Family Environments for Estimates of Heritability and Nonshared Environment in Behavior-Genetic Adoption Studies”, Stoolmiller 1999
- “Does Genetic Variance for Cognitive Abilities Account for Genetic Variance in Educational Achievement and Occupational Status? A Study of Twins Reared Apart and Twins Reared Together”, Lichtenstein & Pedersen 1997
- “Heritability of MMPI Personality Indicators of Psychopathology in Twins Reared Apart”, DiLalla et al 1996
- “IQ Similarity in Twins Reared Apart: Findings and Responses to Critics”, Bouchard 1996b
- “Does Obesity Run in Families Because of Genes? An Adoption Study Using Silhouettes As a Measure of Obesity”, Sørensen & Stunkard 1993
- “A Silent Childhood”, Rymer 1992
- “The Malleability of IQ As Judged from Adoption Studies”, Locurto 1990
- “The Body-Mass Index of Twins Who Have Been Reared Apart”, Stunkard et al 1990
- “Assessment of Effects of Socio-Economic Status on IQ in a Full Cross-Fostering Study”, Capron & Duyme 1989
- “An Adoption Study of Human Obesity”, Stunkard et al 1986
- “Genetic Influences in Criminal Convictions: Evidence from an Adoption Cohort”, Mednick et al 1984
- “The Texas Adoption Project: Adopted Children and Their Intellectual Resemblance to Biological and Adoptive Parents”, Horn 1983
- “Let’s Understand Skodak and Skeels, Finally”, Jensen 1973d
- “A Final Follow-Up Study of One Hundred Adopted Children”, Skodak & Skeels 1949
- “Nature-Nurture and Intelligence”, Leahy 1935
- Miscellaneous
- Link Bibliography
See Also
Links
“Predicting Political Beliefs With Polygenic Scores for Cognitive Performance and Educational Attainment”, Edwards et al 2024
“The Garden of Forking Paths; An Evaluation of Joseph’s ‘A Reevaluation of the 1990 Minnesota Study of Twins Reared Apart IQ Study’”, Bouchard 2023
“Personality Traits, Mental Abilities and Other Individual Differences: Monozygotic Female Twins Raised Apart in South Korea and the United States”, Segal & Hur 2022
“Parent Contributions to the Development of Political Attitudes in Adoptive and Biological Families”, Willoughby et al 2021b
Parent Contributions to the Development of Political Attitudes in Adoptive and Biological Families
“Genetic and Environmental Contributions to IQ in Adoptive and Biological Families With 30-Year-Old Offspring”, Willoughby et al 2021
“Heritability × SES Interaction for IQ: Is It Present in US Adoption Studies?”, Loehlin et al 2021
Heritability × SES Interaction for IQ: Is it Present in US Adoption Studies?
“Does Biology Drive Child Penalties? Evidence from Biological and Adoptive Families”, Kleven et al 2021
Does Biology Drive Child Penalties? Evidence from Biological and Adoptive Families
“Parental Income and Mental Disorders in Children and Adolescents: Prospective Register-Based Study”, Kinge et al 2021
Parental income and mental disorders in children and adolescents: prospective register-based study
“Why Do Wealthy Parents Have Wealthy Children?”, Fagereng et al 2021
“Does Parental Education Influence Child Educational Outcomes? A Developmental Analysis in a Full-Population Sample and Adoptee Design”, Ludeke et al 2021
“The Role of the Shared Environment in College Attainment: An Adoption Study”, Anderson et al 2020b
The role of the shared environment in college attainment: An adoption study
“The Intergenerational Transmission of Early Educational Advantages: New Results Based on an Adoption Design”, Halpern-Manners et al 2020
“Childhood Adoption and Mental Health in Adulthood: The Role of Gene-Environment Correlations and Interactions in the UK Biobank”, Lehto et al 2019
“The Association Between Genetic Predisposition and Parental Socialization: An Examination of Gene-Environment Correlations Using an Adoption-Based Design”, Knoblach et al 2019
“Fullerton Virtual Twin Project: Overview and 2019 Update”, Segal & Niculae 2019
“Genetics, the Rearing Environment, and the Intergenerational Transmission of Divorce: A Swedish National Adoption Study”, Salvatore et al 2018
“Childhood Social Class and Cognitive Aging in the Swedish Adoption/Twin Study of Aging”, Ericsson et al 2017
Childhood social class and cognitive aging in the Swedish Adoption/Twin Study of Aging
“Cross-Generational Transmission from Drug Abuse in Parents to Attention-Deficit/hyperactivity Disorder in Children”, Kendler et al 2016
“The Role of Parenting in the Prediction of Criminal Involvement: Findings from a Nationally Representative Sample of Youth and a Sample of Adopted Youth”, Beaver et al 2015
“Are Adoption Gains on the G Factor? A Meta-Analysis”, Nijenhuis et al 2015
“The Heritability of Alcohol Use Disorders: a Meta-Analysis of Twin and Adoption Studies”, Verhulst et al 2015
The heritability of alcohol use disorders: a meta-analysis of twin and adoption studies
“Family Environment and the Malleability of Cognitive Ability: a Swedish National Home-Reared and Adopted-Away Cosibling Control Study”, Kendler et al 2015
“The Relationship between Parental Depressive Symptoms and Offspring Psychopathology: Evidence from a Children-Of-Twins Study and an Adoption Study”, McAdams et al 2015
“Born in Korea-Adopted Apart: Behavioral Development of Monozygotic Twins Raised in the United States and France”, Segal & Cortez 2014
“Children of Twins Design”, D’Onofrio 2014
“A Closer Look at the Role of Parenting-Related Influences on Verbal Intelligence over the Life Course: Results from an Adoption-Based Research Design”, Beaver et al 2014
“‘Mischievous Responders’ Confound Research On Teens”, Kamenetz 2014
“Inaccurate Estimation of Disparities Due to Mischievous Responders”, Robinson-Cimpian 2014
Inaccurate Estimation of Disparities Due to Mischievous Responders
“Continuity of Genetic and Environmental Influences on Cognition across the Life Span: A Meta-Analysis of Longitudinal Twin and Adoption Studies”, Tucker-Drob & Briley 2014
“Quantitative Genetics in the Postmodern Family of the Donor Sibling Registry”, Lee 2013b
Quantitative Genetics in the Postmodern Family of the Donor Sibling Registry
“The Lost Study: A 1998 Adoption Study of Personality That Found No Genetic Relationship between Birth-Parents and Their 240 Adopted-Away Biological Offspring”, Joseph 2013
“Intelligence: New Findings and Theoretical Developments”, Nisbett et al 2012
“Genetic and Familial Environmental Influences on the Risk for Drug Abuse: a National Swedish Adoption Study”, Kendler et al 2012
“The Causal Effect of Parents' Schooling on Children’s Schooling: A Comparison of Estimation Methods”, Holmlund et al 2011
The Causal Effect of Parents' Schooling on Children’s Schooling: A Comparison of Estimation Methods
“Why Are Children in the Same Family so Different from One Another?”, Plomin & Daniels 2011
Why are children in the same family so different from one another?
“Human Aggression across the Lifespan: Genetic Propensities and Environmental Moderators”, Tuvblad & Baker 2011
Human aggression across the lifespan: genetic propensities and environmental moderators
“Genetic and Environmental Influences on Adult Life Outcomes: Evidence from the Texas Adoption Project”, Loehlin et al 2007
“How Large Are the Effects from Changes in Family Environment? A Study of Korean American Adoptees”, Sacerdote 2007
How Large are the Effects from Changes in Family Environment? A Study of Korean American Adoptees
“Proceeding From Observed Correlation to Causal Inference: The Use of Natural Experiments”, Rutter 2007
Proceeding From Observed Correlation to Causal Inference: The Use of Natural Experiments
“The Mechanism of Emergenesis”, Lykken 2006
“Biology-Environment Interaction and Evocative Biology-Environment Correlation: Contributions of Harsh Discipline and Parental Psychopathology to Problem Adolescent Behaviors”, Riggins-Caspers et al 2003
“The Nature and Nurture of Economic Outcomes”, Sacerdote 2002
“Implications of the Restricted Range of Family Environments for Estimates of Heritability and Nonshared Environment in Behavior-Genetic Adoption Studies”, Stoolmiller 1999
“Does Genetic Variance for Cognitive Abilities Account for Genetic Variance in Educational Achievement and Occupational Status? A Study of Twins Reared Apart and Twins Reared Together”, Lichtenstein & Pedersen 1997
“Heritability of MMPI Personality Indicators of Psychopathology in Twins Reared Apart”, DiLalla et al 1996
Heritability of MMPI personality indicators of psychopathology in twins reared apart
“IQ Similarity in Twins Reared Apart: Findings and Responses to Critics”, Bouchard 1996b
IQ similarity in twins reared apart: Findings and responses to critics
“Does Obesity Run in Families Because of Genes? An Adoption Study Using Silhouettes As a Measure of Obesity”, Sørensen & Stunkard 1993
“A Silent Childhood”, Rymer 1992
“The Malleability of IQ As Judged from Adoption Studies”, Locurto 1990
“The Body-Mass Index of Twins Who Have Been Reared Apart”, Stunkard et al 1990
“Assessment of Effects of Socio-Economic Status on IQ in a Full Cross-Fostering Study”, Capron & Duyme 1989
Assessment of effects of socio-economic status on IQ in a full cross-fostering study
“An Adoption Study of Human Obesity”, Stunkard et al 1986
“Genetic Influences in Criminal Convictions: Evidence from an Adoption Cohort”, Mednick et al 1984
Genetic Influences in Criminal Convictions: Evidence from an Adoption Cohort
“The Texas Adoption Project: Adopted Children and Their Intellectual Resemblance to Biological and Adoptive Parents”, Horn 1983
“Let’s Understand Skodak and Skeels, Finally”, Jensen 1973d
“A Final Follow-Up Study of One Hundred Adopted Children”, Skodak & Skeels 1949
“Nature-Nurture and Intelligence”, Leahy 1935
Miscellaneous
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Link Bibliography
-
2024-edwards.pdf: “Predicting Political Beliefs With Polygenic Scores for Cognitive Performance and Educational Attainment”, Tobias Edwards, Alexandros Giannelis, Emily A. Willoughby, James J. Lee -
2022-segal.pdf: “Personality Traits, Mental Abilities and Other Individual Differences: Monozygotic Female Twins Raised Apart in South Korea and the United States”, Nancy L. Segal, Yoon-Mi Hur -
2021-willoughby.pdf: “Genetic and Environmental Contributions to IQ in Adoptive and Biological Families With 30-Year-Old Offspring”, Emily A. Willoughby, Matt McGue, William G. Iacono, James J. Lee -
2015-beaver.pdf: “The Role of Parenting in the Prediction of Criminal Involvement: Findings from a Nationally Representative Sample of Youth and a Sample of Adopted Youth”, Kevin M. Beaver, Joseph A. Schwartz, Eric J. Connolly, Mohammed Said Al-Ghamdi, Ahmed Nezar Kobeisy -
2014-donofrio.pdf: “Children of Twins Design”, Brian M. D’Onofrio -
2002-sacerdote.pdf: “The Nature and Nurture of Economic Outcomes”, Bruce Sacerdote